
Remember me
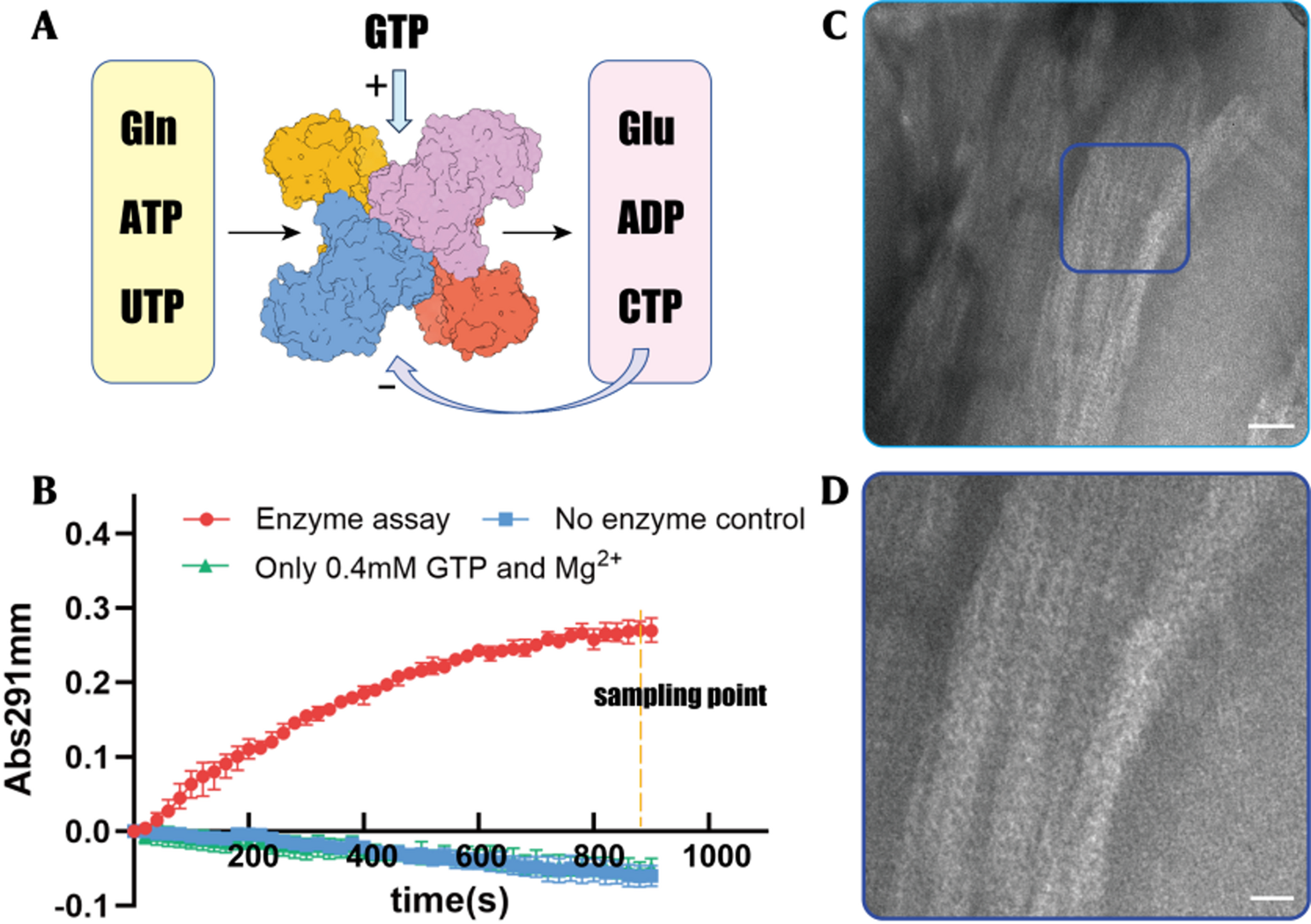


hCTPS1 functions as a homotetramer, with each monomer comprising a glutamine amidotransferase (GAT) domain and an amidoligase (AL) domain. In the presence of GTP, the GAT domain hydrolyzes glutamine (Gln) to produce ammonia, which is transferred to the AL domain to catalyze the conversion of uridine triphosphate (UTP) to cytidine triphosphate (CTP) via an ATP-dependent intermediate, 4-phospho-UTP (4-Pi-UTP) (Fig. 1A). Purified hCTPS1 exhibited high homogeneity and robust catalytic activity in enzyme assays (Fig. 1B).
Fig. 1
Catalytic mechanism and enzymatic reaction of hCTPS1. (A) Reaction scheme of CTP synthase. UTP accepts a phosphoryl group hydrolyzed by ATP and form an intermediate 4-Pi-UTP in the GAT domain (grey box). The ammonia is released from glutamine in the AL domain (yellow box) and then ligated to 4-Pi-UTP to form CTP in GAT domain. (B) Kinetic curves of hCTPS1 enzyme assay in 0.4 mM GTP and control experiments. The X-axis indicates time. The Y-axis indicates the change in absorbance 291 nm. (C-D) Negative-stain EM micrographs of hCTPS1 at the end of enzymatic reaction. Scale bar, 50 nm. (D) is the zoomed-in view of the blue box in (C). The scale bar is 200 nm
In the enzymatic reaction system containing magnesium ions, GTP, and excess Gln, the reaction produced ADP, glutamate (Glu), and CTP. After 1200 s of reaction with 1 mM UTP and 1 mM ATP, the system generated approximately 0.45 mM CTP, achieving a conversion of 45% and a final UTP:CTP ratio of 11:9. Strikingly, cryo-electron microscopy analysis of the reaction solution revealed large, stacked bundles composed of periodically arranged filamentous structures (Fig. 1C, D). Grayscale analysis confirmed the uniformity of these structures, suggesting that hCTPS1 can form filaments and further aggregate into higher-order bundles in CTP-rich environments.
CTP induces hCTPS1 filamentationTo investigate the mechanism underlying hCTPS1 filamentation, we explored the effects of various ligands on its self-assembly. Previous studies have shown that hCTPS1 forms filaments in the presence of substrates (ATP and UTP) and the allosteric regulator GTP (Fig. 2A). Here, we demonstrated that hCTPS1 also forms filaments in solutions containing the enzymatic products CTP, ADP, and Glu (Fig. 2B), indicating that product binding can induce filament assembly.
Fig. 2
Negative staining of hCTPS1 filament. The wild-type hCTPS1 can form filament under several condition with Mg2+. Scale bar, 50 nm. Filaments are indicated by yellow arrows. (A) Negative-staining EM micrographs of hCTPS1 incubated with all substrates (ATP, UTP, GTP and Gln) combinations. (B) Negative staining of hCTPS1 incubated under all product ligands condition. (C) hCTPS1 forms filaments when incubated with CTP and ADP. (D) Incubation of hCTPS1 with CTP alone is sufficient to induce filament polymerization.
By systematically reducing the types of ligands in the reaction system, we identified CTP and ADP as key inducers of hCTPS1 filamentation. Remarkably, hCTPS1 formed filaments in the presence of CTP alone (Fig. 2C) or ADP alone, but not in systems containing only Glu. These results suggest that CTP, this end product, can coexist with hCTPS1 filaments, challenging the prevailing view that CTP triggers filament disassembly.
To further validate this finding, we conducted a time-point sampling in which CTP was added to hCTPS1 and immediately imaged using negative-staining electron microscopy. Filamentous structures were observed within seconds, confirming that CTP rapidly induces hCTPS1 filamentation (Figure S1). This finding contradicts previous models and suggests that CTP-mediated feedback inhibition of hCTPS1 does not involve filament disassembly.
Cryo-EM structure of CTP-bound hCTPS1 filamentTo elucidate the structural basis of hCTPS1 filamentation under product-bound conditions, we resolved the cryo-EM structure of hCTPS1 filaments in the presence of CTP (Figure S2&S3). To stabilize the GAT domain, we included 6-diazo-5-oxo-L-norleucine (DON), a glutamine analog that irreversibly inhibits CTPS activity by covalently binding to the glutamine site. Negative-staining experiments confirmed that DON does not disrupt filament formation.
Cryo-EM analysis revealed clear filamentous structures, and 2D classification highlighted detailed structural features (Fig. 3A). Using 312,573 particles for reconstruction, we achieved a final resolution of 3.3 Å. The structure showed that hCTPS1 filaments consist of helical arrays of tetramers, with each helical unit rising by 105 Å and twisting by approximately 40° (Fig. 3B, C). The tetramers are connected through α-helix-mediated interactions involving residues 346–357 in the GAT domain. Notably, residues H355 and W358, which are critical for filament assembly across species, exhibited continuous electron density in the density map, underscoring their conserved role (Fig. 3D). Each tetramer contained two distinct CTP binding sites, providing insights into the molecular basis of CTP-mediated regulation.
Fig. 3
Cryo-electron microscopy (cryo-EM) analysis and overall structure of hCTPS1 filaments with CTP. (A) Cryo-electron micrograph of hCTPS1 under CTP and DON condition. Several representative 2D class averages of the hCTPS1 filament are selected in view of the RELION-4 on the right. (B) Schematic model of hCTPS1 filament. CTP bound filament model twists 40° between two adjacent tetramers and shifts along the axis 105 Å (C) Cryo-EM structure of hCTPS1 bound to CTP and DON. The central hCTPS1 tetramer is colored by protomer. (D) Zoomed-in view of the top green box in (C), showing the interface of two adjacent tetremers. The density of residues responsible H355 and W358 for the interactions are color red. Zoomed-in view of the below blue box in (C), showing the CTP in the non-canonical site.
Conserved CTP binding modes in hCTPS1 filamentsCTPS is known to undergo product feedback inhibition, where high CTP concentrations induce conformational changes that reduce enzymatic activity. In our structure, two CTP molecules were observed bound to each protomer: one at the interface of three protomers (overlapping with the UTP binding site) and the other within a single protomer (overlapping with the ATP binding site) (Fig. 4A). We designated these as the canonical and non-canonical binding pockets, respectively.
Fig. 4
CTP binding modes in hCTPS1 and comparison of CTP binding pockets among organisms. (A) A single refined hCTPS1 tetramer model from the filament, colored by protomer (pink, goden, blue and red) and nucleotide highlighted in yellow and crimson. Zoomed-in view of the below blue box shows the CTP in the canonical(yellow) and non-canonical(crimson) site, which are respectively occupied by UTP and ATP under substrate state. (B-C) Diversities of CTP binding pockets among organisms. Models of hCTPS1, Drosophila cytidine triphosphate synthase(DmCTPS) and hCTPS2 are colored in pink, blue, and green, respectively, with the corresponding Protein Data Bank (PDB) codes 9vmm, 7dpw, and 7mh1. (B) Canonical CTP binding pocket. CTP is represented by yellow sphere-and-stick models, hydrogen bonds are shown as blue dashed lines, and magnesium ions(Mg2+) represented by green spheres. (C) Comparisons of non-canonical CTP binding sites
In the canonical binding pocket, the triphosphate group of CTP is stabilized by electrostatic and hydrogen-bonding interactions with each protomer. Residue E155 forms salt bridges with the ribose group, while the cytosine base interacts electrostatically with the backbone of residue E155. Sequence alignment revealed that the residues involved in CTP recognition in this pocket are highly conserved in hCTPS2 and Drosophila CTPS (DmCTPS) (Fig. 4B & Figure S5, 6).
In the non-canonical binding pocket, the triphosphate group interacts electrostatically with a loop, while the cytosine group is stabilized by interactions with S21, K319, and F77. These residues are identical in DmCTPS and highly conserved in hCTPS2, except for T250, which differs between hCTPS1 (I250) and hCTPS2 (T250). This difference has been exploited to design selective inhibitors targeting hCTPS1 (Fig. 4C) [40].
Conserved conformational changes in hCTPS1 filamentsLike hCTPS2 and DmCTPS, hCTPS1 filaments exhibit distinct conformations under substrate-bound and product-bound conditions. While the filament assembly interface remains unchanged, the helical parameters of the filaments vary with ligand binding (Fig. 5A & Figure S7).
Fig. 5
Structural comparison between hCTPS1(purple), hCTPS2(blue) and DmCTPS(green) in substrate-state and product-state. The structures of hCTPS1, hCTPS2 and DmCTPS in s-state are colored in pink, powder blue and lime green, with the corresponding Protein Data Bank (PDB) codes 7mgz, 6pk4, and 7dpt. The structural comparison of hCTPS1, hCTPS2 and DmCTPS in p-state are colored in purple, deep sky blue and forest green, with the corresponding Protein Data Bank (PDB) codes 9vmm, 7mh1, and 7dpw. (A) Structural comparison between hCTPS1 tetramer, hCTPS2 tetramer and DmCTPS tetramer in substrate-state and product-state. (B) Structural comparison of AL domain and GAT domain. The wing structure in GAT domain has deviated among the three species, while the conformation of AL domain remains almost unchanged. (C) Rotation axis of AL domain and GAT domain transitioning between two states. (D) Comparison of the interaction interface between adjacent tetramers in filament. H355 and W358 are colored red
Structural alignment of the AL domain (residues 1–270) between substrate-bound (s-state) and product-bound (p-state) hCTPS1 revealed an RMSD of 1.267 Å, with conformational differences localized to the Arch region (residues 45–67) and Loop183–194. The GAT domain exhibited high rigidity, with an RMSD of 0.899 Å for all 249 Cα atoms, indicating stable conformations under different ligand conditions (Fig. 5B).
Further analysis showed that the conformational changes in the hCTPS1 tetramer primarily involve rotational displacement of the GAT domain, while the AL domain remains relatively fixed (Fig. 5C). The filament interface on the GAT domain surface maintains a stable, outward-facing conformation, enabling antiparallel assembly of tetramers. This assembly mode is conserved in hCTPS2 and DmCTPS (Fig. 5D).
In summary, hCTPS1 filaments exhibit conserved conformational changes and assembly interfaces under substrate and product conditions, highlighting the evolutionary conservation of the filamentation regulatory pattern.
Comments (0)