
Remember me
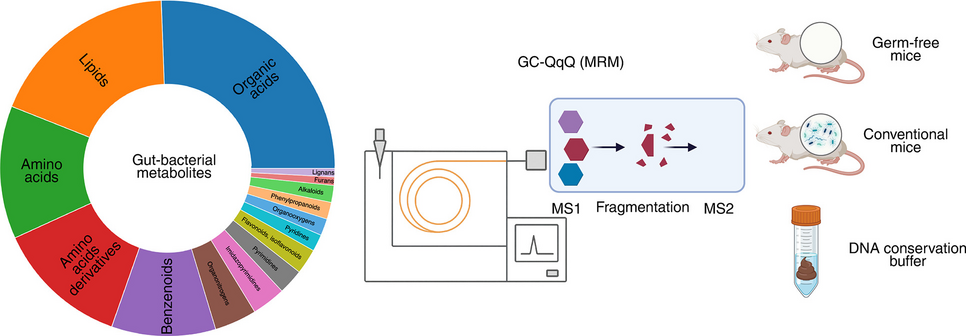

Metabolites included in the method. Asterisks (*) indicate metabolites with matched isotopically labeled internal standards used for absolute quantification. Colors depict chemical classes defined by MiMeDB [31]
Gut microbial metabolite library preparation and metabolite derivatizationTo establish a targeted metabolomics method, we assembled a library of 120 chemical standards of metabolites that are produced by the human gut microbiota and that span multiple metabolite classes as defined by the Microbial Metabolites Database (MiMeDB) [31] (Fig. 1, Table 1, and Supplementary Table S2). The library includes the short-chain fatty acids (SCFAs) acetate, propionate, and butyrate, which are produced during gut bacterial fiber fermentation, serve as important energy substrates for colonocytes and hepatocytes, and can function as modulators of immune regulation [32, 33]. We also included branched-chain fatty acids (BCFAs) [34]; organic acids, such as lactate [35] and succinate [36]; and indoles [37] derived from tryptophan metabolism, all of which are involved in microbial cross-feeding and signaling [38]. Additionally, amino acids [39] and some of their derivatives, including GABA [40], a neurotransmitter linked to gut-brain communication, were part of the library (Fig. 1, Table 1). The selected metabolites range from 60.05 Da (acetic acid) to 412.69 Da (stigmasterol). Further, these compounds have a broad range of boiling points—from low (e.g., short-chain fatty acids) to high (e.g., non-volatile amino acids and dicarboxylic acids, such as succinate and fumarate). In addition, we included isotopically labeled compounds for 52 of the metabolites as internal standards (IS), enabling metabolite quantification (Fig. 1, Table 1, Supplementary Table S3).
To facilitate gas chromatographic separation, we chemically derivatized the metabolites as previously reported by Gu et al. [41]. In brief, we first basified the samples with NaOH to decrease the volatility of organic acids and, hence, enable sample drying under vacuum. Prior to GC-MS/MS analysis, we then performed methoxymation and silylation (using methoxyamine and N-tert-butyldimethylsilyl-N-methyltrifluoroacetamide—MTBSTFA), respectively, to enhance volatility and thermal stability. A key advantage of using MTBSTFA is that the derivatization reaction can be performed at 60 °C, without the requirement of ultrasonication [42]. Moreover, the chemical replacement of polar and reactive functional groups with less polar and more thermally stable groups allows direct injection into the GC-MS instrument without the need of any further extraction steps [43].
Optimization of MRM assaysWe optimized the temperature gradient of the gas chromatography separation for the separation of all metabolites. To this end, all 120 metabolites were injected separately and as a mixture to determine their retention times, which ranged from 2.35 to 24.90 min (Fig. 2a and Table 1) in the final 25-min run.
To optimize the MRM parameters of the triple quadrupole (QqQ) instrument, we then selected one or two precursor ions for each compound and isotopically labelled internal standards. To this aim, we injected each metabolite eight times and ramped the collision energy (CE) of Q2 from 3 to 45 eV to determine specific MS/MS-fragments and the CE resulting in their highest occurrence. Using the Smart Database (GCMSsolution Ver. 4.22), the optimized MRM parameters were managed and together with their respective retention time assembled to the final GC-MS/MS method (Table 1). Confirmation ions were selected to help quantification through improved identification of the peaks upon splitting or retention time shifts.
Table 1 Key parameters for TBDMS-derivatized metabolites and their corresponding isotopically labeled internal standards, including retention time, MRM transitions, collision energy (CE), limit of quantification (LOQ), range of linearity of quantification, and limit of detection (LOD). *IS, internal standard used for the quantification of a specific metabolite.Quantification of metabolitesThe limit of detection (LOD) was defined as the lowest concentration at which the signal for a given metabolite could be reliably distinguished from background noise, corresponding to a signal-to-noise ratio of greater than three. We found an LOD of 0.05 pmol for organic acids, lipids, and lipid-like molecules; 12.34 pmol for short-chain fatty acids (SCFAs); 0.15 pmol for amino acids and their derivatives; and 4.11 pmol for flavonoids, alkaloids, furanoid ligands, and organooxygen compounds (Table 1). For the limit of quantitation (LOQ), we report the concentration for which the signal to noise ratio was greater than 10 (Fig. 2b).
Fig. 2
a Chromatogram and b limit of quantification of 120 gut bacteria-derived metabolites. Colors indicate the MiMeDB metabolite class. c Calibration curves of valeric acid (orange, solid line) corrected with 2H₉-valeric acid as internal standard (orange, dashed line) and of tryptamine without internal standard correction (red, solid line) and with d2-indole-propionic acid for internal standard correction (red, dashed line)
For the absolute quantification of metabolites, we calculated the ratio between the peak area of metabolites and their corresponding isotopically labeled internal standards and compared the value to the corresponding calibration curves. In the absence of a matching internal standard compound, we selected a structurally similar internal standard with close retention time to the analyte (Table 1) to determine metabolite concentrations. This strategy extended the linear range of quantification, as demonstrated for valeric acid and tryptamine, whose calibration curves were corrected using 2H₉-valeric acid and 2H-indole-propionic acid, respectively (Fig. 2c). Following this approach, we achieved a linear quantification range of 0.68 to 500 pmol for organic acids and 1.37 to 1000 pmol for amino acids, branched-chain fatty acids, indole derivatives, and lipid-like molecules. Some compounds, such as short-chain fatty acids (4.11–1000 pmol), ectoine (12.34–1000 µM), propylparaben, piperine, arachidonic acid, erythronic acid, and enterolactone (0.103–1000 pmol), exhibited narrower linear ranges.
To validate the method, we analyzed the certified reference plasma material NIST SRM 1950. Four 100 µL aliquots of the NIST plasma were extracted and derivatized as independent replicates following the developed protocol for sample extraction, derivatization, and GC-MS/MS measurements. Accuracy was evaluated for metabolites for which reference concentrations are available: 15 metabolites with certified values reported by NIST and an additional 9 metabolites with literature values reported by Mandal et al. [44]. These quantified metabolites span four orders of magnitude in concentration (from 0.3 to 2600 µM), allowing the assessment of method accuracy across a broad concentration range in a physiological matrix. For both the NIST-certified set and the literature-reported set of metabolites, the concentrations measured with our method were within the respective reference ranges (Table 2, Supplementary Table S14). These results demonstrate robust analytical accuracy. Further, these analyses suggest using NIST SRM 1950 as external quality-control material for future measurement batches to ensure inter-batch and inter-study comparability.
Table 2 Quantified metabolites in NIST SRM 1950 plasma and comparison to reference concentrations and concentrations recently reported [44]. Standard deviations are based on four independent replicate measurementsAltogether, we established a targeted metabolomics protocol for the quantification of 120 gut-bacteria-produced metabolites using a total of 52 isotopically labeled internal standard compounds. We report LOQ and linear quantification range for each of the metabolites to facilitate the analysis of biological samples.
Quantification of gut microbiota-derived metabolites in the plasma and liverTo demonstrate the utility of the developed targeted metabolomics method for quantifying microbiota-derived metabolites in vivo, we applied it to samples from six conventional (CONV) and six germfree (GF) C57BL/6 mice of mixed sex (Supplementary Table S4). Using the established GC-MS/MS protocol, we detected 86 metabolites in plasma samples in both groups of mice, of which we could quantify 81 (Fig. 3a). Measured concentrations align well with previously reported values, providing an important validation of the established method. For example, we measured a plasma concentration of 519.9 ± 8.2 µM for acetic acid in CONV mice, which aligns with previously reported concentrations of 101 µM and 580 µM (Fig. 3b) [44, 45]. Further, amino acid concentrations in the plasma of GF mice, such as isoleucine (mean, 34.2 ± 4.2 µM; reported range, 29 µM and 50 µM) and aspartic acid (mean, 6.7 ± 0.8 µM; reported range, 5 µM and 11 µM), were within the range of previously reported concentrations (Fig. 3c, d) [46, 47].
Among the 81 quantifiable plasma metabolites, we detected ten solely in the plasma of CONV mice (Fig. 3a, Supplementary Table S5). Furthermore, 40 metabolites showed significant differences (padj < 0.05) between GF and CONV animals, illustrated in the scatter plot (Fig. 3e). Among the metabolites with higher plasma concentration in CONV animals are indole-acetic acid and indole-propionic acid, which are solely produced by microbes but not the host and, hence, should indeed not be detected in germfree animals [20]. Furthermore, microbiota-produced acetic acids, 2-ketoisocaproic acid, and valeric acids are more abundant in the plasma of CONV mice. Altogether, these data demonstrate that our developed method is capable of detecting microbiota-dependent metabolites in plasma samples and that the determined concentrations of specific metabolites are in agreement with previous reports.
Next, we wanted to test our protocol for the measurement of metabolites extracted from solid tissues. To this aim, we extracted metabolites from liver samples from the same animals using the bead-beating protocol previously described [52]. We detected 78 metabolites, 73 of which we could quantify. Measured concentrations also align well with previously reported values. For example, the concentration of glycine (mean, 2.2 ± 0.1 nmol per mg; reported range, 0.6 and 2.7 nmol per mg), glutamic acid (mean, 0.7 ± 0.1 nmol per mg; reported range, 1.2 and 2.1 nmol per mg), and serine (mean, 0.4 nmol ± 0.05 per mg; reported range, 0.32 and 1.00 nmol per mg) in CONV mice (Fig. 3g–i) [50, 51]. Three of the detected metabolites (hippuric acid, suberic acid, and tryptamine) were quantified only in the liver of CONV mice, whereas 12-hydroxystearic acid was only quantifiable in GF mice, which highlights the microbiome dependency of these metabolites. (Fig. 3j, Supplementary Table S6).
Fig. 3
Metabolites quantified in plasma and liver tissues of CONV and GF animals. a Venn diagram representing the number of quantified metabolites in GF and CONV animals plasma samples. b–d Measured and previously reported concentration range of b isoleucine, c aspartic acid, and d acetic acid in plasma samples from CONV mice. e Scatter plot of metabolites quantified in plasma samples of conventional and GF animals. f Venn diagram representing the number of quantified metabolites in GF and CONV animals in liver samples. g–i Measured and reported concentration range of g glycine, h glutamic acid, and i serine in liver samples from CONV mice. j Scatter plot of metabolites quantified in liver samples of CONV and GF animals [46,47,48,49,50,51].
Quantification of gut microbiota-derived metabolites along the intestineSince we measured microbiota-dependent differences in the concentration of specific metabolites in plasma and liver, we next aimed to quantify microbiota-produced metabolites directly in the complex intestinal tract of GF and CONV mice. To this aim, we analyzed the intestinal content of different sections of the intestine (duodenum, jejunum, ileum, cecum, colon, and feces) to quantify metabolites at the site of gut microbial activity. In total, we analyzed 72 samples, including six mouse replicate samples per intestinal section for either group (i.e., CONV and GF). From our panel of 120 targeted metabolites, we could quantify between 63 and 82 in a given intestinal section and mouse group, with site-specific and colonization-dependent differences in detected metabolites (Fig. 4a, Supplementary Table S7-S12).
We first focused our analysis on the cecum because most microbial metabolic activity is expected in this intestinal section [53]. We detected a total of 97 cecal metabolites, 82 of which we could quantify, with 5 and 22 metabolites only quantifiable in the cecum of GF and CONV mice, respectively (Fig. 4b, Supplementary Table S8). For 80 of the 82 quantified metabolites, concentrations remained within the linear range of the method (maximum measured concentration = 987 µM). However, acetic acid and propionic acid levels exceeded this range in three cecal samples from conventional mice. Therefore, we re-measured these samples after doubling the volume of the derivatization solution during sample preparation to dilute the samples. Nevertheless, one acetic acid sample remained above the linear range post-dilution and was therefore excluded from further analysis. Measured concentrations also align well with previously reported values. For example, the concentration of acetic acid (mean, 22.1 ± 3.2 nmol per mg; reported range, 20 and 60 nmol per mg), butyric acid (mean, 24.6 ± 5.7 nmol per mg; reported range, 16 and 48 nmol per mg), and propionic acid (mean, 16,5 ± 0.9 nmol per mg; reported range, 4.00 and 20.00 nmol per mg) in CONV mice was within the previously reported range (Fig. 4c, d, and e) [54,55,56]. Among the 82 quantified metabolites, 57 were significantly different between CONV and GF animals (padj < 0.05), illustrated in the scatter plot (Fig. 4f, Supplementary Table S10) with key microbiota-dependent metabolites labeled, such as acetic acid, valeric acid, propionic acid, indole-propionic acid, isocaproic acid, and isovaleric acid. The elevated levels of these microbiota-associated metabolites in CONV mice, alongside higher amino acid concentrations in GF mice (e.g., proline concentration, GF = 2.2 ± 0.2 nmol/mg vs. CONV = 0.33 ± 0.10 nmol/mg), can be explained by microbial metabolism. Notably, we detected several microbiota-produced SCFA in the cecum, such as butyric acid and propionic acids, that were absent in the plasma of CONV mice (Fig. 3c). This difference is likely due to their rapid consumption by enterocytes in the gut epithelium and the liver.
To further illustrate gut bacterial metabolism, we performed measurements along the intestinal tract (Fig. 4 g, Supplementary Figs. 1, 2, 3, 4 and 5). For example, to illustrate the lack of microbial consumption or transformation of amino acids in the distal colon of GF mice, we show serine concentrations along the gut (Fig. 4 g). Serine concentrations are significantly higher (padj < 0.05) in the distal colon of GF compared to CONV mice, reflecting the absence of a microbiota that consumes serine in the gut GF mice. Contrarily, to demonstrate microbial metabolite production, we show that hydroxyhexanoic and propionic acid concentrations have an increasing gradient along the intestinal tract—particularly enriched in the cecum and colon of CONV mice (Fig. 4 g). These patterns reflect known microbial colonization zones and emphasize region-specific bacterial fermentation.
Fig. 4
Metabolites quantified in intestinal content and tissues of CONV and GF animals. a Number of quantified metabolites across intestinal content and tissues of CONV and GF mice. b Venn diagram representing the number of quantified metabolites in GF and CONV animals in cecum samples. c–e Reported and measured concentration range of c acetic acid, d propionic acid, and e butyric acid in the cecum of GF and CONV animals. f Scatter plot of metabolites quantified in the cecum of CONV and GF animals. The concentration of all 82 quantified metabolites is represented as mean concentrations (nmol per mg) from six mice in each group. g Serine, hydroxyhexanoic acid, and propionic acid concentrations in the intestinal content along the gut of GF and CONV mice. An independent samples t-test was used to compare metabolite levels between the two groups, *padj < 0.05 [53,54,55,56].
Determine method precision and accuracyTo assess the intraday and interday precision of the developed method, we extracted three batches of four replicates of the same sample (i.e., plasma and cecum from both germfree and conventional mice) on three different days. We then measured each sample batch twice on the same day to assess intraday measurement precision and once on the subsequent day to assess interday measurement precision. To illustrate method precision, representative metabolites covering a broad concentration range in plasma and intestinal content sample matrices are displayed (Fig. 5a). For plasma, indole-propionic acid, solely produced by the gut microbiota, and citric acid, produced by microbes and the host, were selected to illustrate different physiological concentrations. For the cecum, butyric acid and glycine are shown. Precision, quantified as %CV, was determined for citric acid: CONV: interday CV = 6.14, intraday CV = 4.77, and GF: interday CV = 5.50%, intraday CV = 5.50%; for indole-propionic acid: CONV interday CV = 6.40%, intraday CV = 7.90%, and for GF: not quantified; for butyric acid: CONV interday CV = 5.65%, intraday CV = 4.27%, and for GF: not quantified; and for glycine: CONV: interday CV = 11.09, intraday CV = 13.14, and for GF: interday CV = 9.64, intraday CV = 7.03. All these values fall within the acceptable range (CV < 20%) defined by current analytical guidelines (Supplementary Table S15) [57]. In total, we could assess the precision for 80 distinct metabolites in all four sample matrices (i.e., plasma and cecum from both germfree and conventional mice), and for all of them, we determined an intraday and interday CV < 20% (Supplementary Table S15).
Quantification of gut microbiota-derived metabolites in DNA/RNA stabilization buffersNext, we tested the capacity of the developed method to detect gut-bacterial metabolites in intestinal content stored in DNA stabilization buffers. To this aim, we pooled and homogenized cecal material from CONV mice and equally distributed four aliquots in three different storage conditions: (i) freshly frozen samples without any additional solution, (ii) samples stored in Invitek stabilization buffer, and (iii) samples stored in OMNIgene gut stabilization buffer. We then froze the samples at −70 °C and analyzed them using our developed GC-MS/MS protocol. We detected a total of 96 compounds in freshly frozen samples, among which we could quantify 82.
For samples stored in Invitek buffer, we could quantify 79 of the 82 metabolites quantified in fresh-frozen samples, with isocaproic acid, 2-ketoisocaproic acid, and isovaleric acid not being detected (Fig. 5a, Supplementary Table S13). All other metabolites showed comparable concentrations between the two sample storage conditions (e.g., butyric acid in freshly frozen samples = 23.9 ± 0.41 nmol/mg compared to 21.35 ± 1.35 nmol/mg in Invitek buffer and alanine in freshly frozen samples = 1.63 ± 0.09 nmol/mg compared to 1.65 ± 0.1 nmol/mg in Invitek buffer).
In contrast, we could quantify all the 82 metabolites in OMNIgut that we also quantified in freshly frozen samples. However, we also found three metabolites that had significantly (padj ≤ 0.5) lower concentration in OMNIgut stabilization buffer compared to fresh-frozen samples, including 2-ketoisocaproic acid, 2-hydroxyisovaleric acid, and isovaleric acid (Fig. 5b, Supplementary Table S13).
Furthermore, we noticed that measured metabolite intensities in Invitek stabilization buffer are generally lower than those from freshly frozen and OMNIgut buffers. The extensive use of internal standard compounds compensates for this for the quantification of metabolites, but the limit of detection is reduced.
Fig. 5
Metabolites quantified in cecum and plasma of CONV and GF mice analyzed in different measurement batches to assess intra- and interday measurement precision and metabolites quantified in cecal samples of conventional animals stored under three different preanalytical conditions. a Box plots of citric acid and indole-propionic acid quantified in the plasma of CONV and GF animals, and butyric acid and glycine quantified in the cecum of CONV and GF animals. The same samples were processed in three independent batches (each containing four identical replicates) on three different days, and each processed batch was measured by GC-MS/MS three times (two times on the same day and once on the subsequent day). b Scatter plot of quantified metabolites in cecum sample stored in Invitek and OMNIgut stabilization buffers and in a freshly frozen sample. The abundances of all 82 quantified metabolites are represented as mean concentrations from four replicates in each group.
Comments (0)