We examined in full spatial detail the occurrence of longitudinal motion in the vibratory responses of the ooC. We showed that longitudinal motion was restricted to the OHC, Deiters’ cells and the tunnel of Corti. It was frequency and level-independent but did depend on the ear’s metabolic state; after death, they disappeared. They were, however, robust against the damage induced by acoustic overstimulation. Data were collected in animals of either sex, we have no indications that the observations are sex-dependent.
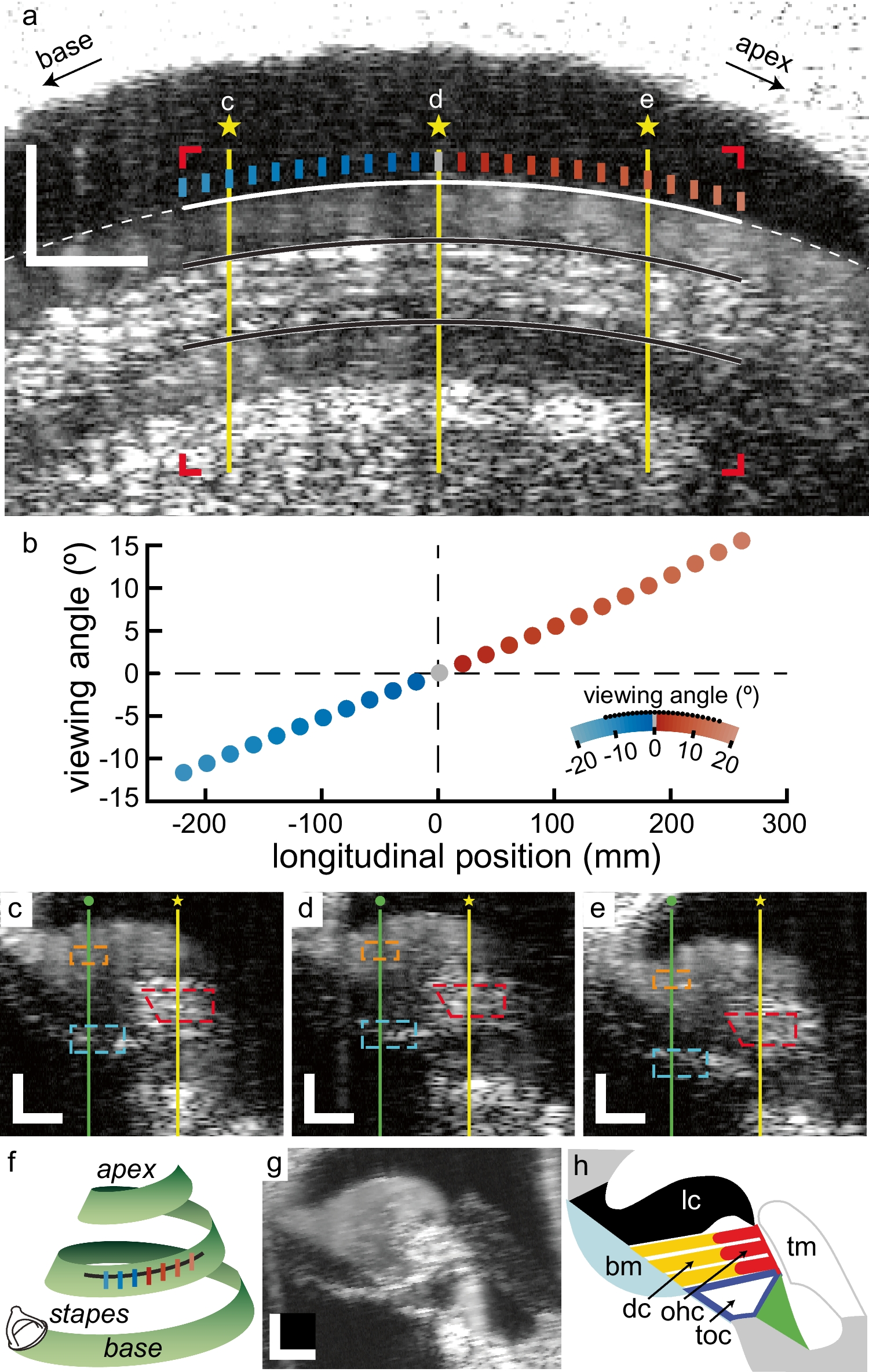
Pivotal in our analysis is that interferometric methods (such as laser Doppler vibrometry or phase-sensitive OCT) only measure the projection of motion onto their measurement beam. With this, the phase of measured motion would flip (changes 180°) when the angle between this motion and the measurement beam changes from acute to obtuse (or vice versa). In effect, vibrations that move towards (away from) the measurement system under acute measurement angles, move away from (towards) the system for obtuse angles. By placing the cochlea under our OCT system such that the direction of longitudinal motion would be near-perpendicular to the (vertical) measurement beam, it is possible to deduce its presence by looking for this 180° phase flip when systematically changing the angle between this motion and the OCT beam.
Rather than rotating the OCT system relative to the cochlea, we used the natural curvature of the cochlea to vary this measurement angle by comparing responses from adjacent tonotopic locations. This introduces several potential errors in the interpretation of the results. First, variation in measurement angle now co-varies with absolute cochlear location, and either can potentially cause phase effects. We have previously shown that the latter is not the cause for the 180° change in phase [8]. Second, and closely related, is the co-variation of measurement angle and tonotopic location. Due to the traveling wave, the response to a tone will have a different phase at two different tonotopic locations, and their difference may be (by chance) close to 180°. We minimized this potential confound by normalizing all phase responses to the location- and frequency specific phase response on the BM, effectively removing traveling wave effects. Moreover, the wavelengths in this region of the cochlea are 1–3 mm [8], which is substantially larger than the longitudinal distance we used to assess the phase flip. So, even if not compensated for, the traveling wave characteristics would cause a phase difference < 180°. The BM is a suitable choice for such a normalization because its vibrations are expected to be dominated by a simple transverse motion [7]. We chose to use BM phase responses that were averaged over a small region, rather than using the phase response of a single pixel located on the BM. Averaging responses has the obvious benefits (e.g., reduce effect of outliers, less selection bias) but may be inaccurate here because of a phenomenon called phase-leakage [16], which can cause large variation in phase. However, we found that for the BM the within-ROI standard deviation of phase was only 0.07 cycle. Moreover, the fact that we found nontrivial and systematic phase difference across positions and along depths in a single beam argues against any significant effects of leakage, as does the systematic reduction and disappearance of the effects with trauma and death, respectively. Nonetheless, we re-analyzed all responses using the phase of single BM-pixels as a reference, were these pixels where selected based on their strong reflectivity (both absolute and relative to their neighbors). This strategy minimizes potential phase-leakage effects. The results of this re-analysis are shown in Online Resource Fig. 3; its similarity to Fig. 5 verifies that phase-leakage indeed did not affect the analysis of our data.
A final potential error comes from the curvature of the cochlea which causes some “skew” [7, 9], in which a single B-scan is not a single “tonotopic plane” of the cochlea. This is akin to the previous potential confound and can be dismissed for the same reason. Given the small angles re. the longitudinal BM orientation (<|15°|), the height of the ooC (< 150 µm from the BM) and the traveling-wave wavelengths, this skew would introduce, at most, an (OHC–BM) phase difference of a few centi-cycles, much less when viewing angle approaches 0°. So, even when only considering the extreme viewing angles, this is clearly not nearly enough to create a 0.5-cycle phase difference when comparing two “oppositely skewed” tonotopic locations.
It is important to distinguish longitudinal motion from longitudinal coupling [7]. Many cochlear modeling studies have postulated an imported role for longitudinal coupling in the ooC (e.g., [17]). Longitudinal motion and longitudinal coupling are different concepts and, in the context of traveling waves, largely independent. For instance, sea waves show significant longitudinal motion in the absence of longitudinal coupling, whereas torsional waves need longitudinal coupling but show no longitudinal motion. Since the current study reports longitudinal motion and contains no tests of longitudinal coupling, we will limit our discussion accordingly. Longitudinal motion has been predicted by several cochlear models (e.g., [18,19,20]) and have been measured in gerbils: in vitro in the tunnel of Corti by tracking motions of the tunnel-crossing nerve fibers [21] and in vivo in the OHC region by combining OCT recordings obtained at two different angles [7, 9]. In agreement with these findings, we systematically observed that the 180° phase flip (indicating longitudinal motion) was restricted to the ooC region occupied by the OHC and Deiters’ cells (i.e., its “core”; Figs., 5, and 7). A rather sharp border excludes the BM from this core region. Sometimes, the region extends into the LC, but we interpret that as an artifact of the alignment/calculation method. That is, within the LC, response phase changes rather suddenly with radial location (e.g., Fig. 4), and small misalignments of the phase patterns from different locations causes “edge-effects” in their difference (i.e., the difference between two identical, but misaligned, patterns increases with the pattern’s gradient, which is largest near edges). This effect is most pronounced when comparing only two measurement angles, and its magnitude depends on the values of compared angles (see Online Resource Fig. 2). Averaging across multiple measurement angles (as was done in Figs. 5 and 7, top row) quickly blurs this edge-effect and reduces the apparent phase-flip area within the LC.
It has been suggested that longitudinal motion within the ooC comes from the hydrodynamics of the cochlea, which causes elliptical particle motion in the direction of the traveling wave at some distance away from the BM [18]. Here, the displacements in the longitudinal direction lag the transverse vibrations by 0.25 cycle, which is observed in experiment [7, 8], our data). Indeed, such elliptical motion is found in all fluid waves; it is an inevitable consequence of incompressibility of the fluid. Purely transverse fluid waves do not exist. In sea waves, for instance, the periodic vertical motion of the water, which causes the visible crests and troughs on the surface, is always accompanied by a periodic horizontal motion of the water along the propagation direction (i.e., longitudinal). Thus, all fluid waves show longitudinal motion, and the ratio of longitudinal to transverse motion in fluid waves is determined by the effective depth of the fluid relative to its wavelength λ, or more accurately, λ/2π [18]. Waves on shallow water (“long waves”) are dominated by longitudinal motion; with increasing depth (or decreasing λ), the relative contribution of transverse motion increases. In the limit of deep water (“short waves”), the magnitudes of transverse and longitudinal motion become equal, and the 0.25-cycle phase difference causes the fluid particles to move in circles. Given these well-known properties of fluid waves, it is entirely expected that cochlear vibration should contain a significant longitudinal component. Its dominance, however, is unexpected from the perspective of classical cochlear models. The key issue is the effective depth of the waves.
In the cochlear partition, the effective depth of the fluid is determined by the vicinity of rigid boundaries. In classical cochlear models (e.g., [18, 22]), these boundaries were taken to be the bony wall of the cochlea. Two aspects of our data speak against this. First, the extreme abruptness of the phase flip reveals a profound dominance of the longitudinal motion. Such dominance can only occur with a very shallow effective depth, i.e., a tiny fraction (≤ 1/20, see [8]) of λ/2π. With a wavelength of ~ 2.5 mm, the distance to the rigid boundaries would be 20 μm or less, whereas the distance of the BM to the bony wall in the phase-flip region is 100 μm or more (Fig. 1). The second observation that speaks against the role of the bony wall as the rigid boundary is the immediate post-mortem disappearance of the phase flip. It is unlikely that the mechanical properties of the bony wall would change within minutes after death. Interestingly, both the close (~ 20 μm) vicinity of the effective rigid boundary and its metabolic dependence are consistent with the assumption that the ooC itself provides the rigid boundaries, and that the degree of rigidity (and with it, the effective “depth”) is under the regulatory control of OHCs. This scenario was previously proposed [7] in connection with both the longitudinal motion and the strong variation of vibration pattern (“mode”) with sound intensity [2, 18].
Alternatively, longitudinal motion could result from OHC-mediated cycle-by-cycle changes in the cross-sectional area of the ooC [19], which is believed to provide a mechanism via which cochlear amplification improves the cochlea’s mechanical response [23, 24]. Necessarily, such area changes displace cortilymph, apparently preferentially in the basal–apical direction [20]. Directionality (i.e., towards the base for area decreases) of this fluid flow may arise from the traveling-wave phase characteristics, creating a peristaltic cortilymph pump. This fluid flow could then provide longitudinal coupling within the cochlea. Such directional, longitudinal coupling also comes from the architecture within the ooC’s core, where the apically slanted phalangeal process of each Deiters’ cell and its basally slanted OHC form an asymmetric Y-shaped element. Our data show that the longitudinal motion is restricted to this region. Apparently only the relatively freestanding bases of the OHC (together with the Deiters’ cells) and the fluids in tunnel of Corti have the degree-of-freedom to move in the longitudinal direction. This suggests a correlation between the cellular architecture and the complex motion within the ooC. A computational model of the mouse cochlea supports the hypothesis that the OHC-mediated sensitivity and frequency selectivity of the mammalian cochlea relies on this Y-shaped geometry [25]. On the other hand, if cycle-by-cycle feedback by OHCs would be the main drive behind the longitudinal motion, it is difficult to understand our observation that the phase flip phenomenon (and thus the longitudinal motion) was unaffected by a degree of overstimulation that significantly reduced cochlear sensitivity. It is conceivable that the overstimulation in this study was did not completely abolish the cycle-by-cycle OHC feedback, and that the remaining OHC was still sufficient to produce a large longitudinal motion. This could be tested in a future study by using pharmacological agents that cause a more drastic elimination of OHC feedback.
In our recordings, we found that longitudinal motions dominate the recorded responses in the live cochlea only; the 0.5-cycle phase flip was not observed postmortem. This seems to favor an active role of OHC as a cause of longitudinal motion, where this activity is present at all stimulus frequencies, levels and even in the (partially) damaged cochlea. Whether this OHC role is via cycle-by-cycle feedback, a (static) creation of “rigid boundaries” within the ooC, or some other, not-yet determined, mechanism remains to be resolved. There can, however, be little doubt about the presence of longitudinal motion within the cochlea. Irrespective if it is an epiphenomenon or arises because of the cochlear mechanics that are involved with sensitive sound transduction (e.g., regulation of mechanical properties, cochlear amplification, longitudinal coupling, stimulus to the inner-hair-cell bundles), its amplitude appears to be larger than that of motion in the transverse/radial plane. This warrants caution when interpreting relative 1D motions that include regions within the ooC that are dominated by the longitudinal response (i.e., exhibit the phase flip). Specifically, measured phase differences between structures may simply reflect a difference in the direction of motion rather than a lead or lag for motions that are parallel to the measurement beam.

Comments (0)