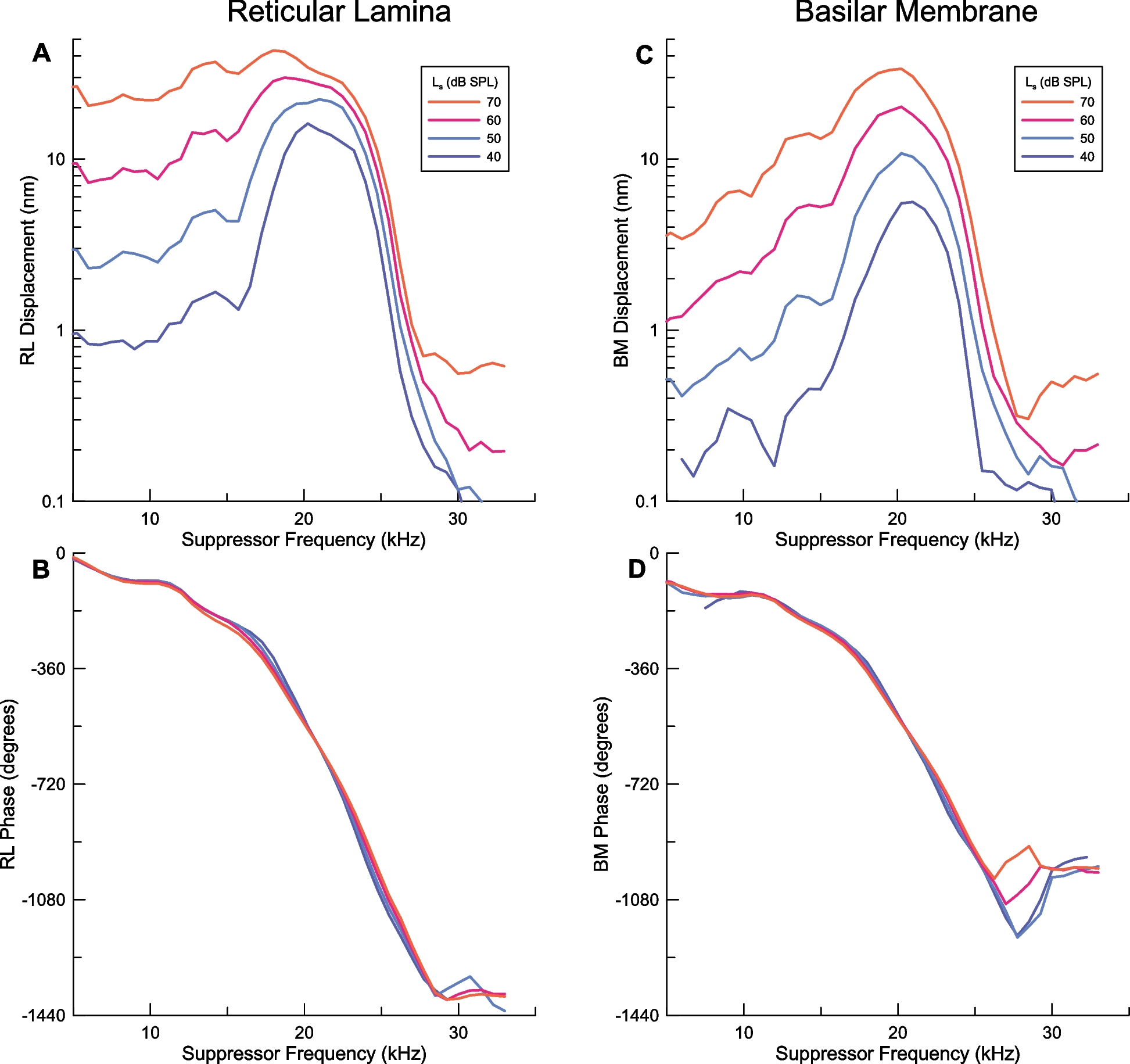
The measurements of vibrations of the RL region and BM reported here show clear evidence for two-tone suppression in the RL region but not the BM for suppressor tones near the ~ 20 kHz local CF acting on fixed probe tones at a frequency two octaves below the CF. Only at suppressor levels of 80 dB SPL is there any hint of nonlinear interaction between the suppressor and the probe in the BM, in some animals. In this situation, the motion of the BM to single-tone excitation does not deviate significantly from linear behavior. Our finding of two-tone suppression in the RL region also adds to the evidence from several labs that active gain in the OHC region is significant for probe tones much lower than the local CF and deviates from linear behavior in other ways. Characterizing this phenomenon was a primary goal of this study.
It should be noted that the position within the organ of Corti where active gain and nonlinearity are observed is somewhat variable between studies. Some studies, including this one, report vibrations of the RL or RL region [7, 10, 12, 15,16,17, 20, 39], while others report that the largest amplified responses are measured in the OHC region [8, 9, 13, 18, 19]. At least part of these differences might be explained by the dramatic changes in the size and submicroscopic structure of hair cells and supporting cells that may constrain motion within the organ of Corti differently, depending on the relative position along the cochlear spiral [40,41,42].
Nonlinearity and Active Gain in the Organ of Corti Below CF
Modern optical measurement techniques revealed the presence of vulnerable gain in the OHC region at frequencies where the BM has been characterized as both passive and linear [7, 9,10,11,12,13,14,15,16,17,18,19,20]. The earliest reports in mice showed level-dependent gain below CF, trending toward linear scaling at the lowest stimulus levels [16, 17]. This trend was also evident in all three turns of the Gerbil cochlea [12]. However, other reports demonstrated that gain below CF scaled almost linearly with stimulus level over a widely varying range of intensity [15, 19, 20, 39]. The gain dropped dramatically postmortem, approaching the gain of the BM in both alive and postmortem states, demonstrating that gain in the OHC region below CF in living animals was active [12, 17, 20]. It was noted later that the gain in the RL region at frequencies well below CF (below 10 kHz) in gerbils was reduced considerably at the highest intensity (80 dB SPL), while linearity was demonstrated in stimulus pressures varying over 5 orders of magnitude, between 20 and 70 dB SPL [15]. While the drop in high-level gain is likely due to saturation of hair cell transduction, it is surprising that the range of stimulus levels where near-linear gain below CF varies so much between experiments, even within the same species.
Level-dependent gain was also reported in so-called hot spots within the organ of Corti [9], but this study used a multitone complex instead of fixed or swept single tones. However, other reports of gain in the OHC region using single-tone stimuli revealed approximately constant gain below CF in the organ of Corti that became more strongly level-dependent during stimulation by a multitone complex [13, 18, 19]. In hindsight, and in view of the results presented here, the multitone stimulus appears to have resulted in mutual suppression in the responses to components of the stimulus, so the single tone results are not equivalent to those obtained with the multitone stimuli and not equivalent to differences in overall stimulus level. These measurements instead provided some of the first evidence of suppression in the motion in the OHC region below CF. More recently, suppression of vibrations below-CF in the OHC region by higher frequency suppressor tones has been demonstrated directly [7, 10]. Additional recent evidence for nonlinearity in the active gain below CF comes from its reduction to near postmortem values in the presence of an intense fixed suppressor tone [19]. The appearance of linear amplification over a range of lower-level stimuli suggests an active quasilinear mechanism at low to moderate stimulus levels. Strimbu and Olson have come to a similar conclusion [19]. However, active gain in the RL region far below CF is reported to saturate and decline toward postmortem passive values for very high stimulus levels in the mouse apex [39]. This shows that the drop in gain to near passive levels does not require either death or irreversible damage to hair cells.
Given that transduction in OHC is demonstratively nonlinear, modeled by Boltzmann functions [43, 44], and that intracellular receptor potentials exhibit harmonic and intermodulation distortion at low to moderate stimulus levels [45], it is puzzling that OHC could produce such near-linear mechanical gain. The situation is reminiscent of operational amplifiers that produce significant gain with distortion reduced through negative feedback. How might such negative feedback happen in the organ of Corti?
The onset of suppression documented in this paper is approximately 40 dB SPL. Likely related is the observation that intermodulation distortion is detected in RL motion for equal level f1, f2 stimuli as low as 40 dB SPL in gerbils [4]. Harmonic distortion and a dc component appear at even lower stimulus levels in the RL region, with consistently lower distortion in BM [46]. While the mildly compressive gain in the OHC region below CF in several reports gives the impression of quasilinear growth, this contrasts with the much stronger compression measured near CF. The appearance of harmonic and intermodulation distortion at low levels, along with two-tone suppression, is much more sensitive measures of deviation from linearity than amplitude scaling. The growth of these sensitive measures of nonlinearity is thus consistent with a mildly compressive extended level range of active gain before strong saturation is measured as seen at the local CF in both the OHC region and BM. This observation indicates that both the active gain and various aspects of nonlinearity seen in the OHC region emerge from the physiology of the outer hair cells and the appearance of these phenomena in BM vibrations represents a filtered version of the nonlinear behavior of the hair cells in the organ of Corti.
Active gain at locations far from the resonant CF place is a recent discovery but has a theoretical basis in hair cell models that generate more mechanical energy than needed to counteract damping [47, 48]. This concept is important in recognizing that cochlear amplification does not necessarily depend on an underlying passive mechanical resonance near the CF place. This is clearly the case for sub-CF active gain demonstrated in experiments. The especially rapid recovery of this gain below CF following furosemide administration [49] shows that it requires only functional outer hair cells and a reasonably normal transepithelial driving force, which is the sum of the endocochlear potential and the potassium ion concentration gradient between the endolymph and perilymph.
There is also a theoretical explanation for the difference in two-tone suppression in the RL region vs. the BM we have observed in measurements. A plausible explanation of our experimental findings is provided by a 3D model of the cochlea showing two-tone suppression of a tone much lower in frequency than the local CF by suppressors near CF in the OHC region but with negligible suppression in the BM at the same location [50]. The difference between suppression in RL vs. BM in the model is because the stiffness of the BM is much greater than the RL. The active force generated by outer hair cells in the model is applied to both the RL and BM, but the much lower stiffness of the RL allows suppression of the hair cell forces to be expressed.
Implications for Otoacoustic Emissions
The evidence of basal suppression reported here is important in its own right as a property of active cochlear mechanics, but it has special relevance to the suppressor method commonly used to separate the SFOAE from the stimulus tone at the same frequency that evokes it [24, 26,27,28,29, 38], a variant of measurements of the otoacoustic phenomenon discovered by Kemp [51].
We believe that our findings address the missing information from prior studies of cochlear mechanics before the modern era of optical methods that can measure vibrations of the BM and the organ of Corti in living animals. We have confirmed that the vibrations of the BM far basal to the peak show the absence of active gain and nonlinearity, with only slight deviation from linearity for 80 dB SPL suppressor tones (Figs. 4 and 5), which likely represents passive coupling from the organ of Corti where two-tone suppression was pronounced. More importantly, our measurements of RL region motion show strong evidence of nonlinearity in the form of two-tone suppression. As noted earlier, nonlinearity at basal locations was required by both the hypothesis that part of the SFOAE arises far basal to the place of the probe tone [23, 25, 26] and by the alternative hypothesis that basal contributions of SFOAEs arise through nonlinear reflection [31, 34]. The other crucial finding, supported by other labs, is that the organ of Corti shows active gain even for probe tones far below the local CF. Furthermore, this amplified probe response is consistently suppressed by a second tone near CF at 40 dB SPL or greater, and the degree of suppression increases with suppressor level. In contrast, we found no evidence that the suppressor tone generated a response at the probe frequency that was not already present in the RL region motion in the absence of the suppressor [34].
There are no conditions in which a suppressor caused an enhancement of the response to the probe, except perhaps for the BM response to 80 dB SPL suppressors in some of our data (Figs. 4 and 5). The suppressor-induced reflection hypothesis was proposed in BM models [32,33,34] but does not appear to have a correlate in the living cochlea for suppressor conditions commonly used to measure SFOAEs [25,26,27,28,29, 38]. Thus, the prediction of these models is not supported by experimental evidence from direct measurements of intracochlear mechanics.
One of the reasons to study SFOAEs is that they represent a measure of otoacoustic emission at a single frequency that can be potentially compared with intracochlear mechanical measures of responses to single tones. The more commonly measured distortion product otoacoustic emissions (DPOAE) suffer from being composed of emissions at intermodulation distortion frequencies, arithmetic combinations of the f1 and f2 stimulus tones. This also includes harmonics of the stimulus frequencies, as well as SFOAE generated by the stimuli.
Determining the relation between intracochlear measurements of mechanics and OAEs measured in the ear canal is a complex problem, primarily because intracochlear measurements such as those we report are typically made at one or a few locations, while OAEs plausibly originate from a broad region of the cochlea despite existing controversy [30, 34].
Given the cochlear frequency-location map, the frequency relationship between the probe and suppressor, the measurement location, and prior SFOAE suppression data, the present results allow us to draw the following important conclusions. First, the basal suppression of the cochlear partition probe response observed in this experiment supports the previous interpretation that SFOAE originates from a broad region basal to the response peak evoked by the probe stimulus [23, 25, 26] This conclusion assumes that two-tone suppression in the RL region relates directly to the changes measured in the ear canal. Our current results are inconsistent with the popular view that SFOAEs are generated through coherent linear reflection (CRF) from the peak region [30]. It must be emphasized that CRF is based on hypothesized random spatial impedance fluctuations in the BM that have never been demonstrated directly in experiments. This hypothesis diverges significantly from the early thinking of Kemp and colleagues that the sources of SFOAEs are more widely dispersed along the cochlea [29, 38].
The other significant finding of our study related to SFOAEs is the absence of evidence for the participation of the BM in generating SFOAE signals or propagating them to the middle ear and to the ear canal. Even for the 5 kHz probe at 60 dB SPL, the suppressor had no significant effect on the levels of the probe response measured in the BM for suppressors of 70 dB SPL or lower (Figs. 4 and 5). This was the case even for suppressors near the frequency of the 5 kHz probe, where the interaction between the suppressor and probe should have been maximized near the place of the probe. Had the BM been the major route of retrograde propagation of the probe signal toward the middle ear, as hypothesized previously [30], then part of the vibration of the BM measured near the 20 kHz place in our data should have changed during the presentation of the suppressor. The OHC region of the organ of Corti appears very likely to be the source of SFOAEs, as vibrations in this region are readily suppressed by external stimuli. Also, the vibrations of the organ of Corti remain large, even far basal to the place of the probe tone, despite decreasing amplitudes of the BM response to the probe tone, likely due to the much greater stiffness of the BM [50].
To connect our intracochlear measurements unambiguously to SFOAEs would require measurements of SFOAEs and intracochlear vibrations in the same animals, in different regions of the cochlea, and in a variety of species during various manipulations such as the application of furosemide [49] or salicylate [52]. Differences in the vibrations of the organ of Corti reported from different labs, species, and cochlear regions suggest that differences in structure may support differing modes of vibration, only some of which may couple effectively to OAEs measured in the ear canal. The hypothesis that suppression in the OHC region invariably leads to evidence of two-tone suppression in the ear canal must be tested directly. But, despite their limitations, OAEs can be used to make measurements of activity as a function of position within the cochlea by varying the frequency or stimuli. Measurements of OAEs can potentially complement intracochlear measurements typically limited to only a few discrete locations.
What Our Results Tell Us About SFOAE Estimation Using the Suppression Method
The suppressor method typically uses two separate measurements of the pressure response in the ear canal: the first with the evoking probe tone by itself and next with the probe tone presented simultaneously with a second (suppressor) tone at a higher level and at a different frequency. The change in the probe tone between the two measurements was referred to as the “residual” by Kemp and colleagues [38, 53] (see also [27, 28]). The change in the ear canal pressure only indicates that part of the SFOAE is suppressed but does not indicate the part of the emission that is not suppressed, a fundamental “blind spot” in OAE measurements using suppressors. The estimate of the SFOAE cannot be accurate unless the suppressor completely eliminates the source of the emission at all cochlear locations where it is generated. But this cannot be known from the ear canal measurements alone. Intracochlear measurements of suppression are beginning to reveal how extensive this problem is.
In the case of the data presented here, the active RL region gain near

Comments (0)