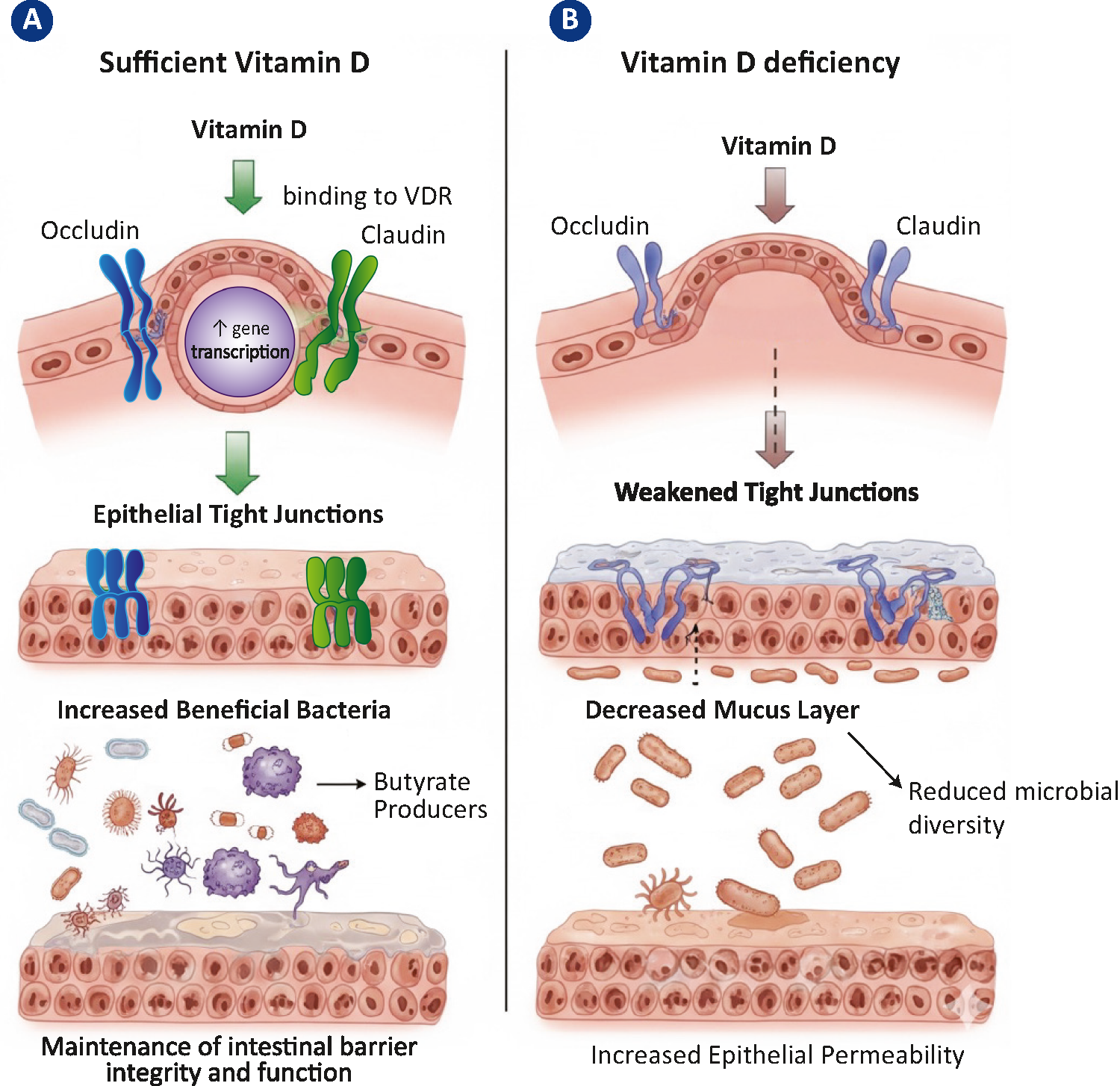
This systematic review represents, to our knowledge, the first attempt to synthesize the existing fNIRS literature across the transdiagnostic spectrum of EDs and obesity. Despite substantial methodological variability, both in terms of experimental paradigms and the clinical populations examined, several neurobiological patterns emerge, which can be interpreted in the context of a dimensional, spectrum-based framework. Across the reviewed studies, the PFC consistently emerges as a central hub of dysfunction, with alterations that may be more appropriately interpreted as domain-dependent rather than diagnosis-dependent. Specifically, convergent alterations were reported in the dlPFC, IFG, and orbitofrontal cortex, regions consistently implicated in cognitive control, value computation, and inhibitory processes [15,16,17, 19, 20, 22,23,24,25,26,27,28, 30, 31]. Importantly, the direction of the effect (hypoactivation versus hyperactivation) was not uniform but varied as a function of the domain engaged. This suggests that altered PFC dynamics in disordered eating should not be interpreted as a simple deficit or excess of control, but rather as an inefficient or context-sensitive modulation of PFC resources. These alterations should be interpreted with caution, as changes in hemodynamic responses do not always correspond to clear behavioural differences – particularly in patients with EDs – and may reflect multiple processes, including compensatory recruitment, variability in task strategies, task-related demands, or physiological confounds.
Four broad functional domains emerged as particularly relevant in this respect: inhibitory control and value-based decision-making, “cold” cognitive load, affective and social-emotional processing, and motor–executive integration.
Concerning inhibitory control and value-based decision making, the most consistent pattern was observed in patients with obesity and with BED. Here, PFC responses were frequently attenuated during paradigms such as Go/No-Go, Stroop interference, and food choice tasks compared to HCs [15,16,17, 22]. This hypoactivation, often accompanied by poorer behavioral performance, aligns with models positing reduced top-down control and impaired integration of reward valuation and executive regulation [40,41,42,43,44,45]. Interestingly, in some subgroup comparisons (e.g., obesity with vs. without BED), individuals with BED exhibited greater dlPFC activation under specific task conditions, such as passive viewing of food images or inhibitory control with appetitive stimuli [15]. Rather than contradicting the broader pattern of PFC inefficiency, this finding may reflect a compensatory attempt to upregulate executive resources in the presence of particularly salient or triggering cues [40]. In this sense, the increased dlPFC recruitment observed in BED does not indicate genuinely enhanced executive control, but rather underscores a domain-dependent inefficiency: greater neural effort is required to manage appetitive cues, and yet this heightened activation does not necessarily translate into improved behavioral outcomes.
A subset of studies employed paradigms that probed executive or language functions without direct involvement of food-related, affective, or social cues. These included verbal fluency tasks (VFT/WFT), working-memory span tests, and classic Stroop interference [16, 25, 27, 28, 30, 31]. Taken together, these studies suggest that executive inefficiency may represent a transdiagnostic feature of EDs and obesity, even when appetitive or emotionally salient stimuli are not present. Verbal fluency tasks consistently pointed to reduced oxygenation in inferior frontal, orbitofrontal, and left frontotemporal cortices in broad ED samples, a pattern in line with diminished engagement of language–executive circuits under neutral cognitive load [27, 28, 30, 31]. Moreover, the prolonged deoxygenation observed in frontal and temporal regions may indicate a slower hemodynamic return to baseline, possibly reflecting either greater cognitive effort or reduced efficiency of recovery mechanisms, and potentially influenced by medication effects [30]. In AN, however, findings were more variable: some studies demonstrated absent task-related modulation or under-recruitment during VFT [27, 28], while others reported no significant differences from HCs [24]. Such inconsistency may reflect a pattern frequently noted in AN, where “cold” cognitive functions do not consistently show clear deficits and can at times appear relatively preserved [46]. These domains may be particularly sensitive to contextual factors such as task demands, age, nutritional status, illness duration, or comorbidities, although the present evidence does not permit firm conclusions.
Further hints come from working-memory tasks, suggesting that executive alterations may manifest more in terms of inefficiency than absolute impairment [46, 47]. In bulimic disorders, for instance, Digit Span Backward elicited similar overall PFC increases compared to controls, but with a more gradual and prolonged activation time-course accompanied by poorer behavioral performance [25].
This profile suggests an inefficient allocation of resources, intended as a greater effort invested without proportional cognitive benefit. In obesity, similar conclusions arise from Stroop interference tasks, where attenuated PFC activation under neutral conflict conditions points to a domain-general deficit in inhibitory control [16].
When tasks targeted affective or social processes, findings varied according to the paradigms employed and the clinical populations examined. In AN, for instance, increased PFC activation was observed in response to attachment-related stimuli, suggesting heightened sensitivity to relational cues [26]. In BN, alterations were linked to subjective experiences of loss of control, with reduced vmPFC and vlPFC engagement during a modified Go/No-Go Task in those reporting more severe loss-of-control symptoms [23]. Similar alterations have been observed in BED, with reduced activation of the left IFG in response to food stimuli [15]. In obesity, by contrast, acute stress paradigms elicited blunted PFC responses, consistent with impaired regulatory capacity under affective challenge [18]. This pattern supports models proposing reduced top-down modulation in stressful or demanding contexts, in line with theories of dysregulated stress responsivity and executive overload[48, 49]. At the same time, increased activation of the left dlPFC has been reported in response to food-related stimuli, pointing to a distinct modulation of PFC circuits by appetitive cues in this group [16, 40]. Taken together, these results suggest that PFC alterations in disordered eating are heterogeneous across affective and social-emotional domains, and their expression depends on the specific processes and clinical groups under investigation.
Finally, studies examining motor-executive coupling provided evidence of reduced neural efficiency in obesity. During dual-task walking and sustained isometric exertion, individuals with obesity exhibited either increased PFC recruitment or attenuated fronto-motor connectivity relative to controls [20, 21]. These results suggest that the integration of motor and cognitive resources requires greater neural effort in obesity, consistent with broader models of reduced efficiency in executive-motor networks. Across these four domains, a tentative distinction emerges between transdiagnostic and condition-specific patterns. Attenuated dlPFC and inferior frontal recruitment during inhibitory control and value-based decision-making represents the most consistent cross-diagnostic signal, observed in obesity, BED, and BN. Reduced frontal activation during verbal fluency tasks across broad ED samples further suggests that executive inefficiency under neutral cognitive load may constitute a shared vulnerability. By contrast, heightened PFC responses to socially and emotionally salient stimuli appear specific to AN, vmPFC and vlPFC alterations tied to loss of control characterise BN, and impaired motor-executive integration under sustained physical demand is most consistently documented in obesity. These disorder-specific profiles suggest that, while the PFC represents a common locus of dysfunction, the functional domain through which this manifests varies systematically across conditions.
4.1 Methodological considerations and limitations
From a methodological standpoint, it is worth noting that the predominance of PFC findings in this review likely reflects not only genuine functional alterations but also the way fNIRS has been applied to date. Most studies employed montages restricted to the PFC surface, a choice motivated by both technical feasibility (reduced interference from hair, flat forehead placement) and by the strong theoretical rationale provided by fMRI and PET studies implicating the PFC in disordered eating. While this has offered important insights into PFC dynamics, it also introduces a coverage bias, leaving other cortical regions relevant to eating pathology, such as insular, parietal, and temporal areas, largely unexplored. Importantly, the absence of findings in these regions should not be interpreted as evidence of lack of involvement but instead entirely reflects a methodological limitation, since these areas were not the focus of investigation in most studies. In addition, a subset of studies investigated clinical samples without control groups. While such designs provided preliminary insights on exercise-related PFC adaptations in obesity, variable neurofeedback effects in BED, and treatment-related changes in BN, the absence of direct comparisons with healthy individuals substantially limits their interpretability. Moreover, fNIRS measurements are inherently sensitive to structural features of the cortex. In conditions like AN, where cortical thinning and brain volume reductions have been consistently documented, changes in scalp-to-cortex distance or cerebrospinal fluid distribution may attenuate the fNIRS signal and add variability[50,51,52,53]. Such factors could have contributed to the relative scarcity of fNIRS research in this population.
At the same time, there are also methodological considerations that support the use of fNIRS in this context. Unlike conventional fMRI, it permits the assessment of cortical activation in ecologically enriched settings and during task conditions that involve movement or embodied interactions, thereby enabling the investigation of neural dynamics that may be difficult to capture in the constrained environment of the MRI scanner [13]. This characteristic is particularly relevant for EDs, where impairments in executive control, reward processing, and sensorimotor integration are often expressed in highly context-dependent ways. The available studies, although still limited in number, illustrate how fNIRS can provide novel insights into these processes, complementing findings from other modalities and contributing to a more nuanced understanding of the neurobiological and cognitive complexity of disordered eating. At the same time, the literature remains sparse and fragmented, with considerable heterogeneity in clinical samples – including age, illness stage, nutritional status, BMI, comorbidities and medication use – experimental paradigms, and analytical approaches. Further limitations include the small sample sizes characterising most studies, the absence of standardised acquisition and analytical pipelines across research groups, and the predominance of cross-sectional designs, which preclude conclusions about the temporal dynamics of cortical alterations across illness stages.
4.2 Opportunities and future directions
Addressing these limitations through larger, methodologically rigorous, and standardized investigations will be essential for consolidating the contribution of fNIRS. Future studies should aim to harmonize experimental paradigms, clinical characterization, and analytical pipelines, thereby improving comparability across research groups [12, 54, 55]. In addition, they should extend fNIRS measurements beyond the PFC to investigate the involvement of other cortical regions, which may contribute to the neurobiology of EDs and obesity. Such efforts would not only enhance the reproducibility of findings but also help to clarify whether observed alterations represent transdiagnostic mechanisms or disorder-specific signatures within the EDs–obesity spectrum.
Importantly, expanding the application of fNIRS in this field has the potential to substantially advance our understanding of the neurobiological mechanisms underlying EDs. A distinctive advantage of this technique lies in its ability to capture cortical activity in ecologically enriched and socially interactive contexts, settings in which many of the core behavioral manifestations of EDs are most evident[12]. In contrast to modalities such as fMRI, which are constrained by artificial experimental conditions, fNIRS allows the investigation of context-dependent neural dynamics with greater ecological validity. Harnessing this potential, while simultaneously improving methodological rigor and sample representativeness, will be crucial for establishing fNIRS as a robust tool to bridge neurobiological findings with clinically relevant phenomena in EDs research. Importantly, beyond its well documented research utility, fNIRS holds specific translational promise. Its portability and tolerance make it a viable tool for real-time monitoring of cortical responses in clinical settings, enabling the tracking of treatment-related neural changes across illness stages. The PFC alterations documented in this review may furthermore serve as potential targets for neuromodulatory interventions, as well as for the development and optimisation of neurofeedback protocols. Establishing reliable fNIRS-based neural markers of dysregulated eating could ultimately contribute to more individualised, mechanism-informed approaches to treatment.

Comments (0)