
Remember me

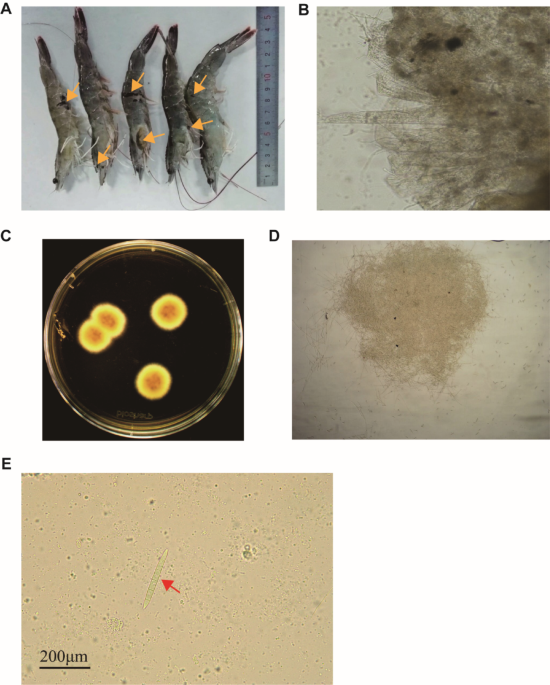

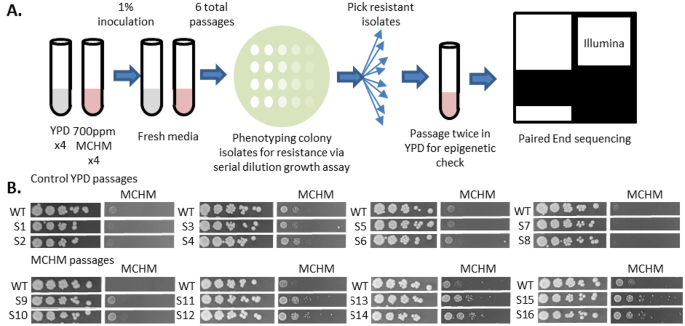
Previous studies using genetic knockouts, transcriptomics, metabolomics, and biochemical assays showed that MCHM resistance in yeast involves complex cellular and genetic networks (Pupo et al. 2019b; Gallagher et al. 2020; Ayers et al. 2020). There is genetic variation in MCHM resistance and YJM789 (a clinical isolate) was more sensitive to MCHM than S96 (S288c background) (Pupo et al. 2019b). To build on this, we used In-Lab Evolution (ILE) to generate resistant strains and identify genetic changes underlying resistance. We evolved four replicates of each strain in YPD with or without 700 ppm MCHM for six passages (Fig. 1A). Two single colonies per biological replicate were isolated after the sixth passage, producing eight control and eight treated strains. Treated YJM789 strains, but not S96, developed increased MCHM resistance, so subsequent analyses focused on YJM789. The YJM789 strains were designated S1–8 as controls and S9–16 as MCHM-treated. All sixteen strains were tested for resistance on YPD with 1000 ppm MCHM.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.In Lab Evolutions produces MCHM resistant yeast strains. A. In Lab Evolutions produce MCHM resistant yeast strains. A Four replicates of YJM789 were grown in YPD ± 700 ppm MCHM, passaged every two days for six passages, and tested for resistance by serial dilution on 1000 ppm MCHM. Resistant isolates were also passaged without MCHM to rule out epigenetic resistance. Two single colonies from each replicate (four control, four treated) were selected for Illumina sequencing. B Resistance was assessed by serial dilution on YPD ± 1000 ppm MCHM. Control strains (S1–S8) and evolved strains (S9–S16) were paired by lineage and tested alongside the YJM789 parent, included on each plate to control for variation in MCHM dosage due to volatility
While the eight control strains showed sensitivity comparable to wildtype, all eight treated strains showed increased resistance relative to wildtype (Fig. 1B). There was plate-to-plate variation in MCHM at 1000ppm, likely due to nearing the solubility limit of the chemical (Sain et al. 2015; Phetxumphou et al. 2016). The wildtype strain was plated on each plate to control for plate-to-plate variation (Fig. 1B).
Genomic analysis of evolved strainsReads from all 16 strains were first mapped to the S288c reference genome, but sequence divergence caused over-calling of SNPs (data not shown). The original YJM789 genome was approximately 5x coverage (Wei et al. 2007). To improve accuracy, we generated a higher-coverage YJM789 reference genome using PacBio sequencing (10 kb average read length, 82× chromosomal coverage, 1154× mitochondrial; Supplemental Table 1). This assembly identified 63,759 SNPs, similar to the ~ 60,000 previously reported (Wei et al. 2007), with ~ 90% intergenic and nonsynonymous variants occurring three times less frequently than synonymous ones (Supplemental Fig. 1). SNPs were distributed genome-wide but enriched near telomeres (Supplemental Fig. 2A). The subtelomeric regions are enriched in nonessential, often duplicated genes that regulate the environmental responses (Brown et al. 2010). The telomeric repeats themselves have heterochromatin that silences the repeats but also extends into the subtelomic regions(Ai et al. 2002). This heterochromatin is established by the Sir protiens and the Mediator tail complex including Med15 are essential for maintaining the heterochomatin (Peng and Zhou 2012).
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Principal Component Analysis of evolved variants in ILE strains by resistance phenotype. Sixteen strains (S1–S16; Supplemental Table 2) were clustered by variant content, with controls (red) and MCHM-resistant strains (blue) labeled as in Fig. 1. The first three principal components are shown: A PC1 vs. PC2, B PC1 vs. PC3, and C PC2 vs. PC3
All 16 strains were sequenced at ~ 25× coverage with Illumina paired end reads. Reads were mapped to S288c and YJM789-specific variants filtered out (Supplemental Table 2). Both control and MCHM-ILE strains carried ~ 1,600 (± 50) mutations across 777 genes. Although MCHM has been linked to DNA damage, we did not observe an increase in mutations in treated cells. Instead, its DNA damage signature appears as G1 arrest (Ayers et al. 2020), which by default reduces DNA replication compared to an asynchronous culture. Across 777 mutated genes, 638 genes were not unique in the MCHM strains and were also in controls. 43 mutated genes were unique to MCHM-ILE strains, but PCA showed no clustering by resistance or lineage (Fig. 2A–C). GO-term analysis revealed growth-related enrichment across all genes (Supplemental Fig. 2D–E), but not among the 43 unique to treated strains, and no copy number variation was detected (Supplemental Table 2).
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Quantitative growth assays of yeast strains with flocculation mutation on MCHM A BY4742 flo1 and flo10 knockouts serially diluted on YPD ± MCHM. B YJM789 and S14 flo1 and flo10 knockouts plated on YPD ± MCHM. C Invasion assay of flo mutants grown ± MCHM for 7 days. D Sedimentation rates of saturated cultures after 5 min without shaking
GO-term analysis of the 777 genes mutated across both control and treated strains revealed enrichment of pathways associated with promoting growth (Supplemental Fig. 2D). Within the control ILE strains, 638 genes were mutated (Supplemental Fig. 2E). After excluding these common mutations, 43 genes remained that were uniquely mutated in the treated strains (Supplemental Table 2). No GO terms were significantly enriched in the treated-only gene set. There was no copy number variation noted in resistant strains.
Filtering MCHM-evolved variants to those involved in coding sequence changesTo assess how mutations contribute to MCHM resistance, we examined the most mutated genes in the 8 ILE strains. FLO1 contained 95 SNPs, including 21 nonsynonymous with predicted moderate effects (Supplemental Table 3), while FLO10 had only six synonymous SNPs. Both encode flocculins that mediate cell adhesion (Rossouw et al. 2015), are regulated by the cell wall integrity pathway (Sariki et al. 2019), and enhance stress resistance via membrane changes (Smukalla et al. 2008). In BY4742 (S288c background), flo1 and flo10 knockouts increased sensitivity to MCHM (Fig. 3A). In YJM789, knockouts had no effect, but in the resistant ILE strain S14, flo1 deletion reduced resistance while flo10 deletion slightly increased it (Fig. 3B). Flo1 likely protects cells by secreting glucose and mannose polysaccharides that limit chemical interactions with the cell wall. Loss of FLO10 did not decrease MCHM resistance in YJM789 suggesting that flocculation is less binary because flo1 knockouts in both BY4742 (BY4742) and YJM789 are more sensitive to MCHM.
Flocculation supports invasive growth in yeast (Sariki et al. 2019) but MCHM inhibited this process (Fig. 3C). Invasive growth is triggered by nutrient starvation but there are significant differences between YJM789 strains. S288c, the lab strain does not flocculate or exhibit invasive growth under any condition. Given that we could not evolve a more resistant S96 (S288c) strain in MCHM and this doesn’t flocculate, we explored the flocculation phenotype in YJM789. Flocculation is induced by the presence of multiple related flocculins on the cell wall. In liquid culture, YJM789 cells readily flocculate and sediment quickly, while flo mutants and the resistant strain S14 remain suspended (Fig. 3D). Therefore, flocculation itself may not be protective against MHCM growth inhibition but there is genetic variation in a multifactorial response to nutrient starvation in which flocculins contribute more than just flocculation. Because flocculation is also linked to invasive growth during nutrient starvation, we investigated other genes mutated in the ILE involved in sugar metabolism. Maltose inhibits flocculation (Stratford 1989) and several maltose transporter paralogs carried missense mutations. MAL31, a permease regulated by Mal13, a transcription factor (Orikasa et al. 2018), is highly divergent between BY4742 and YJM789. A mal31 mutant in BY4742 was sensitive to MCHM (Supplemental Fig. 3A), but knockouts in YJM789 showed very minor effect on MCHM resistance at 650 ppm but not 850 ppm in YPD but there was no change in growth on maltose growth (Supplemental Figs. 3B–C). No matter the genotype, all yeast were more sensitive to MCHM when grown with maltose as the sugar source instead of dextrose.
Analysis of evolved alleles in PDR3.PDR3 was the only gene with nonsynonymous variants in all eight resistant strains (Supplemental Fig. 4A). As a regulator of the pleiotropic drug response via ABC transporters (Delaveau et al. 1994), PDR3 variants were concentrated in the N-terminal 400 amino acids, preserving DNA-binding regions and suggesting truncated alleles in S10, S11, and S14 could remain functional (Supplemental Fig. 4B). We generated 3D structure prediction of Pdr3 (Supplemental Fig. 5A). Structural models showed major C-terminal changes (Supplemental Fig. 5B), but homodimer contact points were unaffected (Supplemental Fig. 4B). Overall, structural predictions did not point to a single domain in Pdr3 mutated in all ILE alleles.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Genetic variation in Pdr3 alleles contributed to the MCHM induced growth inhibition. A Serial dilution growth assays of one of the evolved resistant strains (S11), the YJM789 parent strain, the YJM789 PDR3 knockout strain, the BY4742 wildtype strain, and BY4742 PDR3 knockout were carried out. The growth assays are on increasing concentrations of MCHM from 0 ppm (YPD) to 1000 ppm. B Serial dilution reciprocal hemizygosity growth assays of BY4742 (S288c) and YJM789 hybrid mutants of PDR3. The growth of the wildtype hybrid was compared to hybrids lacking PDR3BY, PDR3YJM, or both (homozygous diploid) on MCHM. evolution that pointed to a reproducible pathway to resistance
To test whether PDR3 truncation confers MCHM resistance, we knocked it out in YJM789 (Fig. 4A). The pdr3 strain showed MCHM resistance similar to S11, which carries the shortest C-terminal truncation of 82 aa (Supplemental Fig. 4A), indicating ILE variants mimic loss of function. Resistance arose in YJM789 but not in BY4742, despite both being passaged in MCHM; four nonsynonymous SNPs differentiate their PDR3 alleles (Pupo et al. 2019a). Hybrid analysis confirmed that PDR3YJM789 conferred to sensitivity, while PDR3BY or loss of both alleles increased tolerance (Fig. 4B). To test if these mutations in PDR3 caused a gain of function, we knocked out PDR3 in S12, with H288D SNP and S14 containing a C-terminal truncation (Supplemental Fig. 6A). Neither knockout changed the growth of the ILE strain. To test if resistance evolved in these strains was specific to MCHM we tested the growth against other chemicals. ILE strains and pdr3 knockouts showed no cross-resistance to vanillin, 4-NQO, β-mercaptoethanol, cycloheximide, or hydroxyurea (Supplemental Fig. 6B).
Binding data in BY4742 (Gera et al. 2022) indicated Pdr3 binds fewer promoters with lower intensity than Pdr1, but deletion of PDR1 enhanced Pdr3 binding at targets including GAC1, SPO24, and RSB1 (Supplemental Fig. 7).We then compared the influence of one paralogous transcription factor on the binding of the other. Binding strength of Pdr1 at different promoters was not altered in the pdr3 knockout (Supplemental Fig. 8A) In contrast, Pdr3 binding at PDR5, SNQ2, RSB1, LDB7, and at its own promoter change in the pdr1 knockout (Supplemental Fig. 8B). We selected nine promoters to examine if there were SNPs in the YJM789 genome that could affect the sequence of known Pdr3 binding sites (Buechel and Pinkett 2024). No SNP differences were detected in Pdr3 binding sites in the promoters (Supplemental Fig. 9). Analysis of Pdr3 motif occurrences between BY4742 and YJM789 genome did not reveal SNPs or position variation between strains (Supplemental Fig. 9). A YJM789 pdr1 mutant was as resistant to MCHM as the pdr3 knockout (Supplemental Fig. 10A), and both mutants showed no survival defects during long-term starvation.
To test whether PDR3 loss altered PDR1 activity, we examined a YJM789 pdr1 mutant, which was just as resistant to MCHM as the pdr3 knockout (Supplemental Fig. 10A), consistent with the absence of PDR1 mutations in ILE strains. Pdr3, Pdr1, and Yrr1 all regulate SNQ2, an ABC transporter whose deletion increases MCHM sensitivity (Cui et al. 1998; Ayers et al. 2020). In contrast, PDR5 deletion caused only mild sensitivity at low MCHM levels (Supplemental Fig. 10B), and previous work showed it is not required for resistance in BY4741 (a strain related to BY4742 with met15 instead of lys2) despite reduced expression during exposure (Gallagher et al. 2020).
Having identified PDR3 mutations affecting MCHM response in all ILE strains, we next examined its regulation through Med15, a Mediator tail component previously linked to MCHM variation between YJM789 and BY4741 (Gallagher et al. 2020) and known to interact with Pdr3 (Shahi et al. 2010). To test whether MED15 loss suppressed pdr3-mediated resistance, we generated double mutants (Fig. 5A). While med15 strains grew slowly but were not MCHM-sensitive, the med15 pdr3 double mutant was as sensitive as the parental strain (Fig. 5A).
IDR predictions of Pdr3 alleles showed no differences between strains (Supplemental Fig. 10C) but genetic interaction with Med15 was critical for response to MCHM. Previous work further uncovered genetic variation in Med15, a component of the Mediator complex, that contributed to variation in yeast MCHM response between YJM789 and BY4741 (Gallagher et al. 2020). As part of the tail in the Mediator complex, Med15 directly interacts with Pdr3 (Shahi et al. 2010). Therefore, in order to test if loss of Med15 could suppress the pdr3 MCHM resistance we generated double mutants (Fig. 5A). The med15 knockouts are slow growing and while not specifically sensitive to MCHM their slow growth is not altered in the presence of MCHM. However, the med15, pdr3 double mutant appeared just as sensitive to MCHM as the parental strain (Fig. 5A). Previous work has shown that zinc levels are increased in MCHM exposure and excess zinc suppresses BY4741 MCHM sensitivity (Pupo et al. 2019a). We found that zinc levels were not increased in the pdr3 mutant upon MCHM exposure, and neither were the levels of other metals tested (Fig. 5B). Dysregulation of the metallome correlated with yeast sensitivity to MCHM. In a QTL analysis, YKE4, a zinc transporter was linked to MCHM resistance. We tested the ability of zinc to suppress MCHM response, but we only noted mild exacerbation of growth inhibition of all YJM789 (Fig. 5C).
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Genetic interaction of Pdr3 and Mediator complex in response to metals A Serial dilution single pdr3, med15, double pdr3, med15 mutants in YJM789 grown on MCHM or rapamycin. B Percent change of elements potassium (K), magnesium (Mg), phosphorus (P), sulfur (S), and zinc (Zn) when yeast from part D were exposed to MCHM. C Serial dilution single pdr3, med15, double pdr3, med15 mutants in YJM789 grown on MCHM with and without excess zinc
During construction of these mutants, we found that the lys2 auxotrophy in YJM789 increased sensitivity to MCHM compared to prototrophs, and pdr3 knockout did not alter growth in the LYS2 background (Fig. 6A). This prompted us to reexamine the influence of Pdr3 on metabolome and specifically amino acid levels when yeast are exposed to MCHM. The metabolome of untreated wildtype and the pdr3 knockout are not well separated on principle component analysis, supporting the hypothesis that under unstressed conditions Pdr3 doesn’t alter the metabolome (Supplemental Fig. 11). However, MCHM alters amino acid levels in wildtype yeast but less so in MCHM treated cells lacking Pdr3 so the profile of pdr3 cells grown in MCHM was closer to that of untreated wildtype and the knockout (Supplemental Fig. 11). We examined how other amino acids and related molecules levels change with MCHM treatment. Intracellular lysine levels remained unchanged despite the altered resistance phenotype (Fig. 6B and Supplemental Figs. 12).
Fig. 6 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Metabolomic analysis of YJM789 and pdr3 during MCHM exposure A. YJM789 and pdr3 mutants with (K+) and without (K-) LYS2 were plated on to YPD with MCHM. B Heatmap of differential metabolite abundance with and without MHCM treatment of YJM789 and pdr3 mutant yeast. Red indicated increased and blue indicated decreased levels of metabolite with MCHM exposure respective to the untreated yeast. C Serial dilution of growth of BY4741 yeast with knockouts of genes encoding glutathione production and recycling, superoxygen dismutases, and catalase. Media was supplemented with glycine, glutamic acid, glutathione, N-acetylcysteine, and ammonia phosphate
Cysteine was consistently reduced in treated strains, which is notable because it is required for glutathione synthesis. Glutathione and related intermediates were also reduced in YJM789 exposed to MCHM but not after PDR3 deletion, consistent with reports that glutathione or oxidized glutathione rescues growth inhibition (Ayers et al. 2020). We tested mutants defective in amino acid transport (gap1), ROS metabolism (sod1, sod2, ctt1) or glutathione metabolism (glr1, gsh1, and gsh2) in the presence of extracellular metabolites. The gap1, glr1, sod1, gsh1 and gsh2 mutants were sensitive to MCHM (Fig. 6C). The growth of all strains could be partially rescued by glycine, GSH, N-acetyl cysteine, or glutamic acid, while ammonium phosphate restored growth in nearly all strains but not pdr5, gap1 or sod1 (Fig. 6C). These findings support the idea that glutathione metabolism is central to MCHM response, while excess nitrogen from most sources can largely bypass its effects on growth.
To explore glutathione metabolism in the response to MCHM exposure we reexamined the ILE genomes. S12 had mutations in GEX2 which transports glutathione, while Sam3 transports S-adenosylmethionine (SAM) and polyamines (Ames and Lovell 2011). SAM is the precursor to methionine that is then converted to cysteine, the rate-limiting step in glutathione synthesis (Walvekar and Laxman 2019). Both BY4742 gex2 and sam3 mutants were sensitive to MCHM (Fig. 7A). In YJM789, the gex2 mutant was slightly more sensitive, whereas sam3 showed no difference from the parent strain (Fig. 7B). Mutations in SAM3 and GEX2 could contribute to increased resistance of the ILE S12 strain only when in combination with mutated PDR3 or the mutations could have arisen independently as passenger mutations. However, two other ILE strains (S10 and S14) also had nonsynonymous mutations in GEX2 (Supplemental Table 3). SAM3 also had mutations in S9, S13, and S16 strains. S12 and S13 were single colonies isolated from the same tube and SAM3 had identical mutations. The other mutations of SAM3 and GEX2 arose independently, suggesting that they contributed to MCHM resistance. Pdr1 and Pdr3 are not known to regulate glutathione directly and based on the metabolism glutathione is elevated in pdr3 mutant compared to wildtype (Supplemental Fig. 12). The S12 ILE strain has a single nonsynonymous H288D SNP in PDR3 (Supplemental 4B). Perturbing glutathione homeostasis altered the cellular response to MCHM, consistent with previous work showing that exogenous glutathione suppresses growth inhibition in yeast. (Ayers et al. 2020). Similar effects were observed in mammalian (HEK293T) cells and Xenopus embryos, where excess glutathione, but not ascorbic acid, rescued viability after MCHM exposure (Perfetto et al. 2021).
Fig. 7 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Quantitative growth assays of yeast strains with transporter genes knocked out on different concentrations of MCHM A BY4742 knockouts of sam3 and gex2 were serial diluted and plated on YPD plates with and without MCHM. B YJM789 and S12 ILE knockouts of sam3 and gex2 were serial diluted and plated on YPD plates with and without MCHM
Comments (0)