Wickerhamomyces anomalus, which was previously known as Pichia anomala, is among the cosmopolitan yeasts that have a broad environmental distribution. This yeast species is commonly found in fermented food or beverage products, plant surfaces, gut of insects, soil, and water environments (Lenka et al. 2021). This indicates that W. anomalus could thrive in an environment with variable nutrient availability and stress conditions (Padilla et al. 2018; Atitallah et al. 2020; Fathiah et al. 2023). W. anomalus BT3 was originally isolated from alcoholic beverages, our previous study revealed that this strain possesses notable tolerance of a broad range of environmental stresses, including [e.g., high temperature, oxidative stress, osmotic stress] (Lenka et al. 2021; Fathiah et al. 2023). These traits are highly desirable for bioremediation applications, where microorganisms are often exposed to unstable and harsh environmental conditions. Moreover, the origin of BT3 from a food-associated source suggests that it is unlikely to pose safety concerns, which is further supported by the fact that the W. anomalus species are regarded as Generally Recognized As Safe (GRAS). This enhances the potential of BT3 as a secure and sustainable bioremediation agent, especially in scenarios where environmental or regulatory safety is a priority.
Based on the screening of cadmium tolerant strain, BT3 was regarded as the cadmium-tolerant yeast. Indeed, the MIC value of BT3 against CdCl2 toxicity was found in the concentration of > 1000 µM. Very limited studies are available regarding cadmium-tolerant yeast, however, yeast S. cerevisiae is among the tolerant ones. Previous studies revealed the MIC of CdCl2 of S. cerevisiae is 50 µM (Trilisenko et al. 2017). Interestingly, other yeasts of P. hampshirensis 4Aer and C. tropicalis were reported as metal-resistant, which had MIC > 2000 µM and > 800 µM Cd2+, respectively (Rehman and Anjum 2011; Zhang et al. 2019). Interestingly, the spot assay revealed that W. anomalus strains exhibited relatively reduced growth at 40 µM cadmium compared to 120 µM and higher concentrations. This unexpected pattern suggests that low-level cadmium exposure may not be sufficient to fully activate the yeast’s metal stress response systems, leading to greater sensitivity. In contrast, exposure to higher cadmium concentrations might induce protective mechanisms such as metal efflux, chelation, or antioxidant responses, resulting in improved survival. This reflects an induced stress response or cross-protection mechanism that enhances tolerance when exposed to a threshold level of stress. Similar phenomena have been reported in yeast S. cerevisiae, where sublethal temperature stress levels can produce stronger inhibitory effects than more extreme exposures due to insufficient adaptation or incomplete activation of defence pathways (Markiewicz-Potoczny and Lydall 2016).
Cadmium exposure did not significantly affect the growth of yeast BT3 grown in a medium containing up to 200 μm CdCl2. Indeed, BT3 could enter the log phase within 24 h of incubation under those CdCl2 treatments. Higher concentrations of CdCl2 exposure, however, prolonged the yeast lag phase, as 96–168 h were required to enter the log phase. Previous studies reported that the occurrence of stress could extend the length of the lag phase as yeast cells may pause their division cycle in the lag phase to adapt towards Cd toxicity before resuming growth and cell cycle checkpoints. Such phenomenon was found in Aspergillus aculeatinus and A. sclerotiicarbonarius when exposed to glycerol-induced osmotic stress (Hamill et al. 2020). Lag phase extension has also been correlated with complex systems that require cells to develop specific mechanisms for carefully regulating the levels of intracellular metal ions, thereby maintaining metal balance and avoiding toxicity (Rolfe et al. 2012).
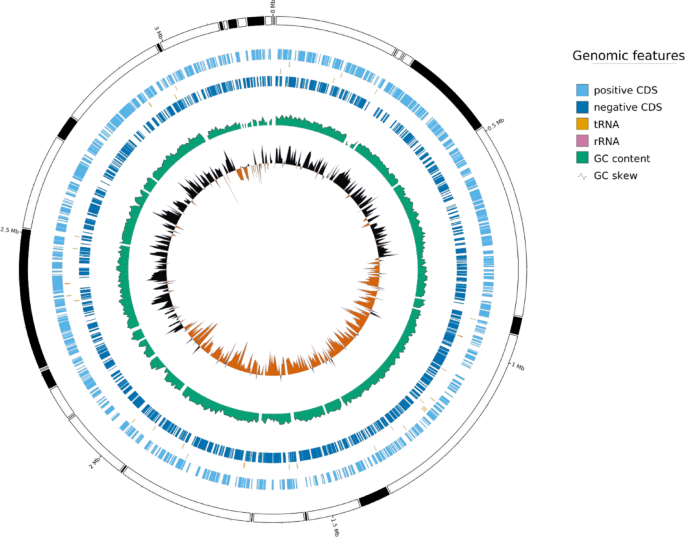
Whole-genome sequencing of W. anomalus BT3 yielded high-quality genomic data, as indicated by Q20 (98.14%) and Q30 (93.75%) scores, which exceed the 80% threshold (Yang et al. 2019). Additionally, the robust 50x depth of coverage confirms the overall validity of the genomic dataset. BT3 has a GC percentage of 35.03%, this is relatively similar to that of reference yeast W. anomalus NRRL Y-366-8 with 34.5% GC. However, other yeast genera of Wickerhamomyces were observed to have various GC content such as W. ciferrii NRRL Y-1031 F-60-10 (ASM31348v1), W. silvicola (ASM3057079v1), W. queroliae (ASM3067425v1), and W. bovis (ASM370794v3) which have 30.5, 32, 39.5, 40.5% GC, respectively.
Through genome sequence data analysis, we were able to identify some genes that were potentially involved in the cadmium-tolerant phenotype of strain BT3. YCF1 gene, which encodes an ATP-binding cassette (ABC) transporter, were observed in the BT3 genomes that could promote cadmium sequestration activity. Cadmium-bound glutathione in the cytosol will be actively transported by YCF1 into the vacuole using ATP hydrolysis for detoxification (Wei et al. 2014; Bickers et al. 2021). Cadmium exposure directly induces oxidative stress, which causes lipid peroxidation and cytoplasmic membrane damage, as reported in yeast S. cerevisiae (Huang et al. 2018). It is, therefore, the oxidative stress response pathway plays a crucial role in coping with cadmium-induced oxidative stress. Genes belonging to the oxidative stress response pathway were identified in the BT3 genome, including SOD1, SOD2,GSH2, PRX1, TRXA, and GLRX. SOD1/2 encode enzyme superoxide dismutase (SOD) with Cu-Zn and Fe-Mn cofactors, respectively. Both genes and the corresponding enzyme activities increased following cadmium exposure in yeast S. cerevisiae and P. kudriavzevii, whereas their genetic mutation resulted in a cadmium-sensitive phenotype (Adamis et al. 2004; Li et al. 2018). GSH2 expresses glutathione synthetase, which is involved in glutathione synthesis. Glutathione could chelate cadmium to form GSH-Cd, which can be further transported to the vacuole by YCF1 for vacuolar detoxication. It is worth noting that a deletion of the GSH2 gene in yeast S. cerevisiae (gsh2Δ) leads to a cadmium-sensitive phenotype (Dai et al. 2024). PRX1, TRXA and GLRX encode peroxiredoxin, thioredoxin and glutaredoxin which are involved in maintaining cellular redox balance and scavenging reactive oxygen species following oxidative stress conditions (Okada et al. 2014). Previous studies reported the involvement of signaling molecules such as nitric oxide in yeast for oxidative stress response pathways that lead to the (Astuti et al. 2016, 2018a).
Genetic regulation in yeast following cadmium exposure may involve transcription factors CAD1 and CUP2, of which the corresponding genes were found in the genome sequence of BT3. Previous studies reported that the expression of CAD1 and CUP2 is induced following cadmium exposure in yeast (Shiraishi et al. 2000). CAD1 is an AP-1-like transcription factor that binds to the promoter of oxidative stress response gene for glutathione synthesis in yeast S. cerevisiae (Kumawat and Tomar 2022). On the other hand, the CUP2 Copper-binding transcription factor activates transcription of the metallothionein genes CUP1-1 and CUP1-2 which are involved in cadmium chelation (Dong et al. 2013). The presence of gene regulatory properties of CAD1 and CUP2 indicates that BT3 manages cadmium-stress exposure through several mechanisms. Indeed, previous reports have noted that to mitigate the severe effects of heavy metal toxicity, organisms must maintain metal homeostasis, evolve mechanisms to control intracellular metal ion levels, or develop effective metal detoxification pathways. Among the complex sensing and regulatory mechanisms, the activity of CAD1 and YCF1 is an essential mode of action in yeast for coping with cadmium toxicity (Gomes et al. 2002; Dai et al. 2024).In line with these findings, the cadmium-resistant BT3 strain may also exhibit cross-resistance to a range of other stressors, including copper, oxidative stress, and nutrient deprivation. The presence of metal-responsive genes and stress-related transcriptional regulators in its genome suggests a broader adaptive capacity that extends beyond cadmium tolerance. The BT3 strain is also likely to possess resistance to copper and a variety of other environmental stress conditions, owing to the presence and predicted functional roles of CUP2, CUP1-1, and CUP1-2. In S.cerevisiae, these genes (particularly CUP1-1 and CUP1-2) have been shown to be strongly induced in strains that demonstrate resilience not only to copper, but also to other toxic agents such as silver ions (Terzioğlu et al. 2020), resinous propolis(Demir-Yılmaz et al. 2025) and nutritional stress like starvation (Arslan et al. 2018). Interestingly, the strain adapted to starvation stress also displayed an extended chronological lifespan along with enhanced copper resistance, implying a possible intersection between stress response systems and longevity mechanisms. In support of this, several independent studies have reported that increased tolerance to copper and the accumulation of cadmium are frequently associated with improved survival under toxic metal exposure, longevity promotion, and active regulation of metallothionein-related pathways (Pabis et al. 2021).
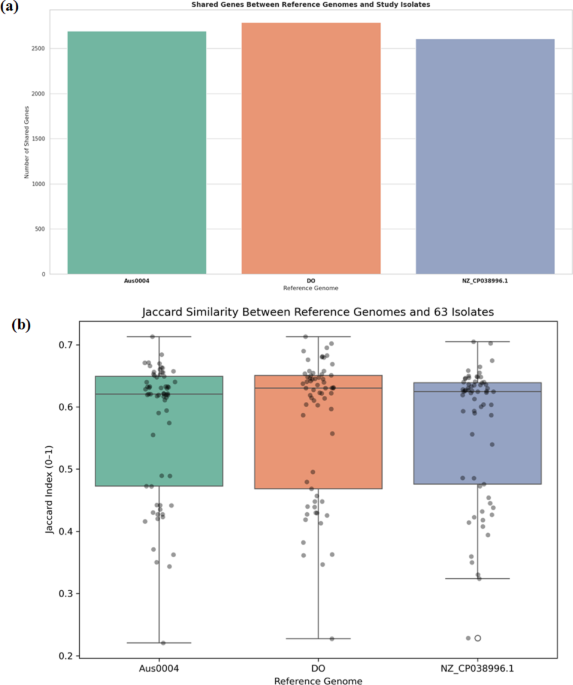
Comparative genome analysis revealed relatively similar genomic features of BT3 to reference genomes, especially to that of yeast from the genera of Wickerhamomyces. In addition to 2212 gene clusters shared among yeast species of W. anomalus NRRL Y-366-8, W. ciferrii NRRL Y-1031 F-60-10, P. kudriavzevii CBS573, K. pastoris ATCC 28,485, P. membranifaciens NRRL Y-2026, S. cerevisiae S288c, S. pombe 972 h-, and C. albicans 101, BT3 shared 702 additional gene cluster with yeast W. ciferrii and W. anomalus, and another 475 gene clusters were shared only with W. anomalus. Those gene clusters account for more than half of the total gene clusters identified in BT3 (6103 gene clusters). Interestingly, 212 gene clusters were identified as being specific to the genome of BT3. Further gene ontology data analysis confirmed that these unique 121 gene clusters were mostly assigned to GO biological process of transmembrane transport (65 genes), GO cellular component of integral component of membrane (7 genes) and GO molecular function of dimethylsulfide monooxygenase activity (33 genes). The large number of unique transmembrane transport-related genes indicates a variability of BT3 in sensing signals and nutrition in the environment. Indeed, transporter protein would be essential for importing nutrients, exporting waste products, and maintaining cellular homeostasis in various conditions (Shao et al. 2025). In addition, dimethylsulfide monooxygenase is an enzyme catalyzing the oxidation of dimethyl sulfide to dimethyl sulfoxide, a crucial step in the sulfur cycle. This enzyme is involved in sulfur metabolism, which, in turn, affects the metabolism pathway in fermentative yeast to create distinct aromas in fermentation products (Kinzurik et al. 2020)(Kinzurik et al. 2020). It is worth noting that BT3 was originally isolated from traditional fermented beverages, which have unique aroma features.
Phylogenetic tree analysis positioned W. anomalus BT3 close to W. anomalus NRRL Y-366-8, and W. ciferrii NRRL Y-1031 F-60-10. Furthermore, W. anomalus BT3 exhibited the most significant gene family expansion, with 103 expanded clusters, among the compared yeast species. This substantial expansion, which contrasts with a smaller contraction of 12 gene clusters, indicates that W. anomalus BT3 has the highest rate of genetic diversification in this group. Novel functions contributing to its enhanced cadmium tolerance could predominantly arise from this genetic diversification.
Our study presents new information about the genome characterization of non-conventional yeast W. anomalus BT3 in relation to its cadmium-tolerant phenotype. The cadmium-tolerant phenotype of BT3 is supported by the genomic properties that are potentially involved in the cadmium sequestration and oxidative stress response mechanism. Some potential key genes responsible for the cadmium-tolerant properties of BT3 have been identified, including YCF1 and CAD1, which are involved in transporting the Cd-GSH complex to the vacuole for metal homeostasis and detoxification. Thus, further studies are needed to explore the molecular mode of action of BT3 in coping with cadmium toxicity involving these genes attributes.



Comments (0)