
Remember me




In early reports, oligosaccharide moieties in the ALOS were found to be diverse, with two major oligosaccharides OSG and OSN, determined by NMR and MS analysis [17, 18]. In the current work, to clearly explore the structural diversity of ALOS, the fully deacylated lipopolysaccharides from A. muciniphila were characterized using LC-Q-TOF mass spectrometry. Mass spectra in negative-ion modes and collision-induced dissociation (CID) fragmentation experiments were optimized for each selected parent ion that provided extensive information to elucidate sequences of oligosaccharide chains (OS). As shown in the ion chromatogram, the oligosaccharides showed a high heterogeneity resulting from the presence of fourteen distinct OS species at m/z 869.7, m/z 909.7, m/z 950.3, m/z 990.3, m/z 1104.3, m/z 1144.3, m/z 1185.3, m/z 1225.3, m/z 1224.4, m/z 1266.4, m/z 1306.4, m/z 1347.5, m/z 1387.4 and m/z 1622.0 (Fig. 1A–C). Six of these variants (m/z 869.7, m/z 909.7, m/z 950.3, m/z 990.3, m/z 1224.4, and m/z 1622.0) were also observed in our previous report based on in-source fragmentation HPLC–ESI–MS analysis [14]. Based on the doubly-charged ions detected in the mass range of m/z 850–1650, the reported monosaccharide composition in ALOS, and the structure of OSG and OSN [17, 18], the sugar sequences in OSA-OSN was elucidated. For example, the oligosaccharide OSF was detected at m/z 1144.3572 [M-2H]2− with a retention time of tR = 3.03 min, which was deduced to be the dodecasaccharide composed of three 3-deoxy-D-manno−2-oct-ulopyranosonic acid moieties (Kdo-1, 2, and 3), two fucoses, one heptose, four hexosamines, two hexoses, and two phosphoric acid moieties (Supplementary Table 1). The CID MS/MS spectrum of OSF indicated the presence of daughter ions at m/z 1034.3 (−2), 1789.6, 1569.5, 1349.5, 939.2, 719.1, and 499.0, as shown in Fig. 2A, B. The occurrence of a fragment ion at m/z 1034.3 (−2) was due to the loss of one Kdo moiety from the double-charged parent ion at m/z 1144.3572 (−2). Two paired fragment ions (m/z 1789.6 and 499.0; 1569.5 and 939.2) were formed due to the cleavage of the glycosidic bond between GlcNP and Kdo-1, between Kdo-1 and Kdo-2 from OSF, respectively. The ions at m/z 1569.5 and 719.1 were derived from the daughter ions at m/z 1789.6 and 939.2 due to loss of one Kdo, respectively. By the similar way, the structural sequences of oligosaccharides OSE and OSH-M corresponding to m/z 1104.3 (−2), 1185.3 (−2), 1225.3 (−2), 1266.4 (−2), 1306.4 (−2), 1347.5 (−2), and 1387.4 (−2) were determined (Fig. 2C, Supplementary Fig. 1A-M, and Supplementary Table 1). Of note, the inner core oligosaccharide of ALOS was found to be composed of three Kdo, one fucose, one heptose, and one hexosamine. The structures for oligosaccharides (OSA-OSF and OSH-OSM) need to be further assigned by sophisticated purification and NMR data elucidation. The diverse oligosaccharides reflect the structural diversity of LOS in the commensal A. muciniphila (Scheme 1), which is deduced to play essential roles in the regulation of bacterial colonization and immune regulation.
Fig. 1
Analysis of core oligosaccharide in the LOS from A. muciniphila by LC-Q/TOF–MS in the negative ion mode. A The total ion chromatogram (TIC) recorded for the core oligosaccharide from A. muciniphila. B Double-charged molecular ion peaks detected in the LC-Q/TOF-MS spectrum. C The extracted ion chromatograms (EICs) of different core oligosaccharide species from A. muciniphila.
Fig. 2
The CID MS/MS spectrum of the oligosaccharide OSF and proposed structures of oligosaccharides detected in A. muciniphila. A CID MS/MS spectrum from the double-charged mother ion at m/z 1144.3. B A proposed fragmentation pattern for OSF. C The proposed structural sequences of the core oligosaccharide from A. muciniphila sketched according to the symbology of monosaccharide residues.“Hex”, hexose; “Hep”, heptose; “Fuc”, fucose; “HexN”, hexosamine; “P”, phosphate; “Kdo”, 3-deoxy-D-manno-2-oct-ulopyranosonic acid, respectively.
Scheme1.
Proposed distribution of lipooligosaccharides on the outer membrane of A. muciniphila
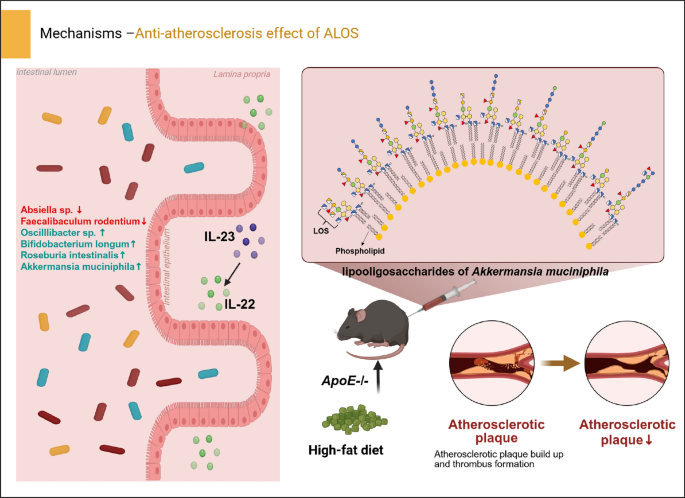
2.2 ALOS inhibits atherosclerosis in ApoE −/− miceIn previous studies, A. muciniphila was reported to attenuate atherosclerotic lesions by reducing metabolic endotoxemia-induced inflammation through restoration of the gut barrier[19]. Interleukin (IL)−23 and its downstream target IL-22 have been reported to limit the development of atherosclerosis by inhibiting the pro-atherosclerotic microbiota[20]. Based on the beneficial effect of A. muciniphila on atherosclerosis and the immunological trait of ALOS, we explored the possible role of ALOS in atherosclerosis. ApoE−/− mice fed with the high-fat diet were intraperitoneally treated with ALOS or vehicle (PBS) for a period of 8 weeks (Fig. 3A). By the end of treatment, plasma lipid levels were significantly improved by ALOS. In comparison with vehicle-treated ApoE−/− mice, reductions in the levels of total cholesterol (TC; 55% and 29%), low-density lipoprotein cholesterol (LDL-C; 44% and 38.9%), total triglycerides (T-TG; 40% and 32%) were achieved by ALOS at doses of 0.2 and 0.5 mg kg−1, respectively (Fig. 3B). The levels of very-low-density lipoprotein (VLDL) and oxidized LDL (ox-LDL) were also significantly reduced by ALOS in a concentration-dependent manner (Fig. 3C–D). The high-fat diet induced formation of atherosclerotic lesions, as demonstrated by hematoxylin and eosin staining, Oil Red O staining, and Masson staining of aortic root regions (Fig. 3E). Compared to Mod, treatment with ALOS substantially reduced the aortic root lesion (Fig. 3E). After 12 weeks of high-fat diet, the accumulation of lipid in the whole aorta was shown in Fig. 3F. Treatment with ALOS substantially reduced the lesion area, necrotic core burden, and the content of collagen (Fig. 3E–G). In addition, ALOS also reduced the lipid accumulation in the liver (Fig. 3H). Based on these results, ALOS showed promising anti-atherosclerosis efficacy.
Fig. 3
ALOS attenuates atherosclerosis in ApoE−/− mice. A Experimental design showing groups and durations (n = 8). ALOS: lipooligosaccharides derived from A.muciniphila. ALOS-L: 0.2 mg/kg ALOS treated- ApoE−/−. ALOS-H: 0.5 mg/kg ALOS treated- ApoE−/−. B Plasma lipid parameters (TC, T-TG, LDL-C, and HDL-C). C Plasma very low-density lipoprotein (VLDL). D Oxidized low-density lipoprotein (ox-LDL). E Representative H&E staining, Oil Red O staining, and Masson staining of cross-sections of aortic roots and quantitative data of plaque area, ecrotic core area, and collagen-positive area (scale bar, 100 μm, n = 4). F Lipid content of the whole aorta visualized by staining with Oil Red O. G The quantitative data of Oil Red O-positive staining area; n = 4 mice for each group. H Representative Oil Red O staining of liver (scale bar, 100 μm, n = 4). *P < 0.05; **P < 0.01. One-way ANOVA (with Tukey's HSD) test in B–H.
2.3 ALOS ameliorates gut barrier function and promotes IL-22 productionEndotoxemia caused by the increased intestinal epithelial permeability due to long-term high-fat diet is recognized as one of the pathogenesis mechanisms underlying the development of atherosclerosis[21, 22]. Compared with atherosclerosis model mice, ALOS-treated mice showed significantly lower plasma LPS levels and high-sensitivity C-reactive protein (hs-CRP) (Fig. 4A–B). Furthermore, ALOS decreased the plasma levels of proatherogenic cytokines, such as IL-6, TNF-α, and increased the levels of anti-inflammatory IL-10 (Fig. 4C–D, and F), while the level of IL-1β remained unaltered (Fig. 4E). The intestinal epithelium serves as the first defense barrier of the gastrointestinal tract and its compromise directly induces inflammation. AB-PAS staining revealed significant reductions of intestinal tissue in goblet cell count in the atherosclerosis model group (Fig. 4G), while ALOS supplementation mitigated the damage to epithelial integrity. In our earlier study on metabolic syndrome, it has been confirmed that ALOS has a weak TLR4 agonist and can elicit the IL-23/IL-22 axis, which is essential for maintaining the integrity of the intestinal barrier and modulation of gut microbiota[17]. We hypothesize that IL-23/IL-22 might be one of the mechanism by which ALOS suppress atherosclerosis. In ApoE−/− mice, the plasma level of IL-22 showed a marked decrease compared to that of normal mice. Compared to atherosclerosis model mice, a high dose of ALOS treatment significantly increased the plasma levels of IL-23 and IL-22 (Fig. 4H–I). The production of intestinal IL-22 was also reduced in the mice with atherosclerosis, while ALOS treatment significantly increased the protein levels of IL-22, as shown by immunofluorescent staining and WB (Fig. 4J–K). These results suggest that the repair of ALOS on the intestinal barrier in atherosclerotic mice is mechanistically related to the activation of the IL-23/IL-22 pathway.
Fig. 4
ALOS ameliorates gut barrier function and promotes IL-22 production. A C-reactive protein (hs-CRP). B LPS. C Tumour necrosis factor-alpha (TNF-α). D Interleukin-6 (IL-6). E Interleukin 1β (IL-1β). F Interleukin 10 (IL-10). G Representative AB-PAS staining of ileum pathological sections (scale bar, 100 μm, n = 4). H Plasma IL-23. I Plasma IL-22. J Representative immunofluorescent staining of IL-22 in the ileum. n = 4 mice for each group. Scale bar, 20 μm. K Western blotting of the IL-22 and β-actin levels in the ileum tissue of ApoE−/− mice. *P < 0.05; **P < 0.01. One-way ANOVA (with Tukey's HSD) test in A–I. Unpaired t-test in J–K.
2.4 ALOS reshapes the structure of the intestinal microbiotaGiven the role of intestinal IL-22 in regulating antimicrobial peptides production and shaping gut microbiota composition[23,24,25,26], we next examined the influence of ALOS on the composition of intestinal microbiota in ApoE−/− mice by the high-throughput sequencing of 16S rRNA in the cecal content. A total of 1256267 sequences were obtained after denoising with DADA2, averaging 89,733 sequences in each sample. Treatment with ALOS did not affect the α-diversity of the bacterial community, as indicated by the Chao1 index and Dominance index, suggesting that ALOS did not change the richness and diversity of the gut microbiome (Fig. 5A–B). Principal coordinates analysis (PCoA) analysis revealed a distinct clustering of gut microbiota composition between the two groups (Fig. 5C), and the UPGMA clustering analysis also showed a clear separation between the model and ALOS groups (Fig. 5D). To further investigate specific alterations of bacterial communities, we compared the relative abundance of the major microbial taxa at the phylum, genus, and species levels. Firmicutes, Bacteroidetes, and Verrucomicrobia were the dominant bacterial phyla in both groups (Fig. 5E). The Firmicutes to Bacteroidetes (F/B) ratio was significantly lower in the ALOS-treated groups (3.18) compared to the mod groups (9.53), corresponding to a 66.63% decrease (Fig. 5E). Meanwhile, ALOS mainly up-regulated the abundance of the genus of Akkermansia, Roseburia, Oscillibacter, and Butyricimonas and decreased the abundance of pathogenic bacteria belonging to Helicobacter and Mucispirillum (Fig. 5F). The linear discriminant analysis effect size (LEfSe) identified 15 operational taxonomic units (OTUs) altered by ALOS, with a significant increase in Bifidobacterium, Robinsoniella, and a decrease in Absiella (Fig. 5G). As to the abundance of species, ALOS treatment decreased the relative abundance of Limosilactobacillus reuteri, and Faecalibaculum rodentium, while enriching the abundance of Bifidobacterium longum, Roseburia intestinalis, Phocaeicola vulgatus, Phocaeicola faecalis, and Akkermansia muciniphila (Fig. 5H–N). These shifts conferred metabolically benefits. F. rodentium, which was decreased by ALOS, contributes to the susceptibility of metabolic disorders by suppressing segmented filamentous bacteria and reducing intestinal Th17 cells [27]. Conversely, B. longum enriched by ALOS exhibited beneficial effects against metabolic disorders by regulating the mRNA expression of renin-angiotensin system factors and ameliorating lipid and glucose levels [28]. R. intestinalis, a promising next-generation probiotic, reinforced barrier function and suppressed pro-inflammatory signaling via butyrate production[29].
Fig. 5
ALOS reshapes the structure of the intestinal microbiota. A The Chao 1 index. B The dominance index. C Principal coordinate analysis. D UniFrac distance-based unweighted pair-group method with arithmetic (UPGMA) means analysis. The shorter the branch length between samples, the higher the similarity between the two samples. E Relative abundance of gut microbiota at the phylum level. F Relative abundance of gut microbiota at the generic level. G LEfSe analysis of microbiota. H–N The relative abundance of differentially abundant flora identified at different taxonomic levels. O Spearman’s correlation analyses with serum biochemical parameters and IL‐22 levels. P Predict microbial function on the METACYC database. PWY-5676 (acetyl-CoA fermentation to butanoate II), BIOTIN-BIOSYNTHESIS-PWY (biotin biosynthesis I), FOLSYN-PWY (superpathway of tetrahydrofolate biosynthesis and salvage), PWY0-1296 (purine ribonucleosides degradation), OANTIGEN-PWY (O-antigen building blocks biosynthesis (E. coli)), PWY-6385 (peptidoglycan biosynthesis III (mycobacteria)), PWY4FS-7 (phosphatidylglycerol biosynthesis I (plastidic)), PWY4FS-8 (phosphatidylglycerol biosynthesis II (non-plastidic)). *P < 0.05; **P < 0.01. Unpaired t-test in A–B, H–J, L–O. Mann–Whitney U test in K, P.
To further investigate the correlation of the key bacteria induced by ALOS and the atherosclerosis-associated symptoms, a Spearman correlation analysis was conducted. As a result, L. reuteri, Absiella sp., and F. rodentium, which were decreased by ALOS, were positively correlated with the parameters of atherosclerosis, while the ALOS-enriched bacteria, such as Oscilllibacter sp., B. longum, R. intestinalis, P. vulgatus, and P. faecalis, were negatively correlated with the symptoms of atherosclerosis (Fig. 5O). To predict the functional alteration in the gut microbiota after treatment, the PICRUSt2 analysis was employed. The results revealed that ALOS upregulated the metabolic pathways of PWY-5676 (acetyl-CoA fermentation to butanoate II), BIOTIN-BIOSYNTHESIS-PWY (biotin biosynthesis I) and FOLSYN-PWY (superpathway of tetrahydrofolate biosynthesis and salvage), which could benefit for ameliorating atherosclerosis [30,31,32], while downregulated the pathways, such as PWY0-1296 (purine ribonucleosides degradation), OANTIGEN-PWY (O-antigen building blocks biosynthesis (E. coli)), PWY-6385 (peptidoglycan biosynthesis III (mycobacteria)), PWY4FS-7(phosphatidylglycerol biosynthesis I (plastidic)), PWY4FS-8(phosphatidylglycerol biosynthesis II (non-plastidic)) (Fig. 5P). Collectively, these data suggest that ALOS treatment partially regulated the structure and functions of gut microbiota in ApoE−/− mice. In conclusion, ALOS treatment ameliorates the dysbiosis of gut microbiota in mice with atherosclerosis, thereby contributing to the alleviation of atherosclerotic injuries.
Comments (0)