
Remember me
The following section presents an integrative thematic synthesis derived from the literature reviewed on immuno-inflammatory mechanisms in pain chronification. Although this work is a narrative review and does not follow systematic review protocols, the organization of content reflects recurring mechanistic domains identified across high-quality studies. These themes are grouped under psychosocial, cellular, molecular, and therapeutic dimensions that converge in the transition from acute to chronic pain. This narrative structure offers a coherent interpretation of complex neuroimmune interactions supported by experimental and clinical evidence.
Neuroimmune Priming by Psychosocial and Perioperative Risk FactorsAlthough comprehensive large-scale studies examining pain chronification remain limited, emerging evidence has identified multiple risk factors that predispose individuals to pain chronification, highlighting that the immune system operates within a broader neuroendocrine-immunological framework [14, 15]. In both pediatric and adult populations, psychological factors significantly influence pain outcomes. Analysis of existing data suggests that individuals may become predisposed to chronic pain through cognitive and emotional pathways, anticipatory anxiety, and fear-avoidance behaviors [16]. The experience of trauma, whether accompanied by pain or not, represents a significant risk factor for chronic pain development, with the social environment playing a crucial modulatory role [17]. In pediatric and adolescent populations, high levels of parental catastrophizing, anticipatory anxiety, and maladaptive coping have been linked to increased postoperative pain and delayed recovery [18,19,20]. These psychological stressors are known to modulate hypothalamic–pituitary–adrenal (HPA) axis activity, alter glucocorticoid sensitivity, and bias T cell polarization toward pro-inflammatory T helper (Th) 1 and Th17 profiles, thereby enhancing glial reactivity after injury [21]. Perioperative clinical factors—such as high baseline pain intensity, pre-existing functional disability, certain high-risk surgical procedures (e.g., spinal fusion, thoracotomy), and large surgical wounds—can increase local tissue inflammation and amplify the recruitment of neutrophils, macrophages, and T lymphocytes [22,23,24,25]. This can lead to neuroimmune priming of the dorsal horn and dorsal root ganglia (DRG), making central sensitization more likely. These risk factors do not operate alone but interact synergistically, creating vulnerability windows during critical developmental periods, particularly in adolescence when neural plasticity is heightened. The interplay between these psychosocial and perioperative factors and immune–neural mechanisms is summarized in Table 1, which links each risk factor to its potential pathway for promoting neuroimmune priming and pain chronification.
Table 1 Psychosocial and perioperative risk factors for chronic pain: neuroimmune implicationsNeuroimmune Interactions in Pain PerceptionThe subjective experience of pain emerges from complex neuroendocrine-immune interactions that can be conceptualized through three dimensions: sensory-discriminative (location and intensity), motivational-affective (emotional response), and cognitive-evaluative (interpretation and meaning) [26]. Within this framework, immune cells and their secreted proteins play crucial roles in modulating nociceptive processing at multiple levels of the nervous system.
Cellular Mediators of PainPain and inflammation are intrinsically linked, particularly in experimental models where controlled tissue injury triggers predictable inflammatory cascades. These models consistently demonstrate elevated levels of prostaglandins, histamine, and platelet-activating factor (PAF), and a coordinated release of cytokines including interleukin-1 (IL-1), IL-6, TNF-α, and interferons [27, 28]. The persistent upregulation of these cytokines supports the presence of local inflammation due to tissue injury and has been shown to activate microglial cells. Once activated, microglia further contribute to the production of pro-inflammatory cytokines such as IL-1, IL-6, and TNF-α, thereby maintaining or amplifying pain states. However, cytokine release is not restricted to the immune system alone; cells of the nervous system also express cytokines. For instance, brain-derived neurotrophic factor (BDNF) is synthesized by neurons and can interact with other neurons within this pro-inflammatory environment, contributing to pain amplification [29]. BDNF triggers intracellular second messenger pathways with neuronal proliferative potential [30, 31].
Another molecule of interest is nerve growth factor (NGF), initially recognized for its role in embryogenesis [32]. In both the postnatal and adult periods, NGF sensitizes nociceptors (particularly those in the DRG) and promotes mast cell activation, leading to sustained pain [33]. Therapeutic inhibition of NGF using monoclonal antibodies, such as tanezumab, has shown promise in experimental models, particularly in chronic pain associated with osteoarthritis, highlighting its pivotal role in nociceptive modulation [34,35,36].
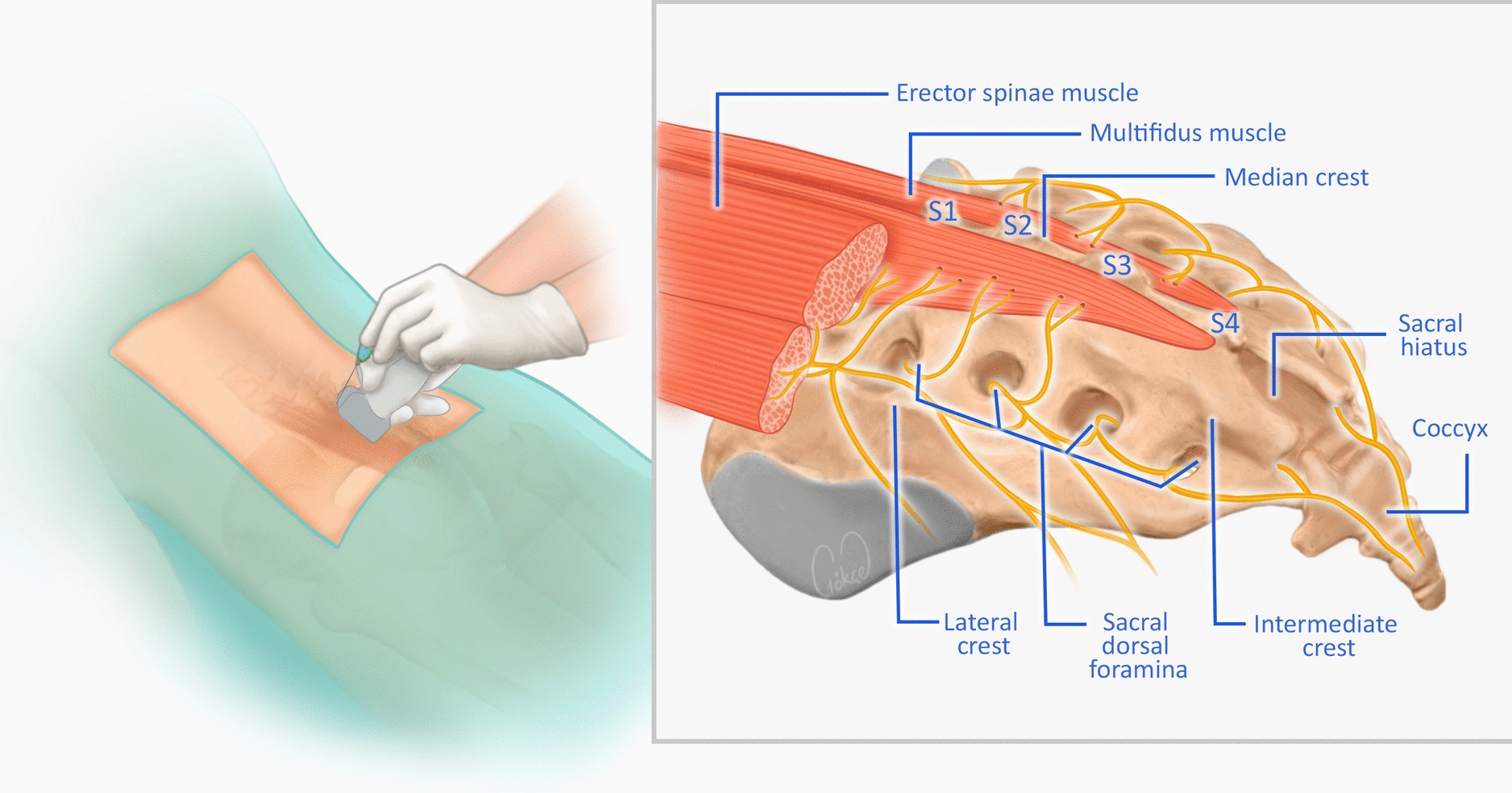
The bidirectional interactions between microglia, neurons, and immune mediators in sustaining or resolving pain are depicted in Fig. 1.
Fig. 1 The alt text for this image may have been generated using AI.
The alt text for this image may have been generated using AI.Microglial–neuronal interactions in pain modulation. Under homeostatic conditions, microglia support neuronal function through trophic factors, phagocytosis, and debris clearance. Inflammatory stimuli shift microglia toward a pro-inflammatory phenotype, releasing cytokines that promote neuronal degeneration and amplify pain. Neurons and mast cells contribute by releasing neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF), further modulating microglial activity and sustaining inflammation. Conversely, anti-inflammatory microglia can counteract these effects, promoting tissue repair and pain resolution
It has been confirmed that local inflammation plays a more critical role than systemic inflammation in the persistence of chronic pain [14, 37]. Systemic suppression of TNF-α has not demonstrated consistent effectiveness in alleviating pain. However, localized interventions (such as intraspinal administration of anti–TNF-α agents) have shown the ability to modulate pain, particularly in models of post-osteoarthritic pain [14].
Several studies have investigated the therapeutic effect of eliminating pro-inflammatory cytokines involved in pain generation [14, 38,39,40]. Interestingly, while the rationale suggested that blocking these cytokines would alleviate pain, the outcomes were not as straightforward. In some cases, systemic cytokine inhibition failed to produce the anticipated analgesic effect, suggesting that local cytokine activity, rather than their systemic presence, may be more relevant in the pathophysiology of chronic pain [14]. These findings highlight the need to reevaluate the spatial and functional dynamics of cytokine signaling in nociceptive pathways.
Similarly, local release of calcitonin gene-related peptide (CGRP) is more closely associated with chronic migraine than its systemic presence [15]. For this reason, it is much more difficult to analyze localized inflammation in humans, and most studies are therefore conducted in rats, where serial biopsies can be performed to evaluate which substances increase more than others and at what stages in a given tissue or injured area [41]. All these cytokines released during inflammation act on second messengers such as nuclear factor kappa-light-chain-enhancer of activated b cells (NF-kB), mitogen-activated protein kinases (MAPKs), and Janus kinases (JAKs), all of which are activated to amplify the inflammatory signal and promote the release of more cytokines such as IL-1, IL-6, and TNF-α [42,43,44]. These, in turn, induce sensitization of neurons in the dorsal horn of the spinal cord and the dorsal root ganglion, increasing pain hypersensitivity. This initial inflammatory phase is driven by the innate immune system, including neutrophils, macrophages, and dendritic cells, among others [45].
From this initial mechanism, it has been found that localized inflammation activates T lymphocytes (T cells), specifically Th1 cells, which secrete pro-inflammatory interleukins such as IFN-γ and IL-2 [46]. These cytokines reactivate macrophages to release more cytokines and promote proliferation. Another T cell involved in pain is the Th17 lymphocyte, along with its IL-17A, which has been implicated in pain [47, 48]. It appears that there is a prior signaling event, as a specific class II major histocompatibility complex (MHC), haplotype DQB1*03:02, has been associated with a higher risk of chronic pain, in concordance with the presence of CD4+ T cells and Th17 cells in the dorsal root ganglion [49,50,51]. In cases of disc herniation with nerve root compression, radicular pain has shown a CD4+ T cell component, predominantly expressing IL-2 and IL-4, possibly activating a B cell-mediated humoral response [52].
On the other hand, T cells that predominantly secrete IL-10 and IL-4, such as CD4+ T cells or Th2 cells, appear to play a protective role against the development of chronic pain [53]. However, this duality of functions remains under debate.
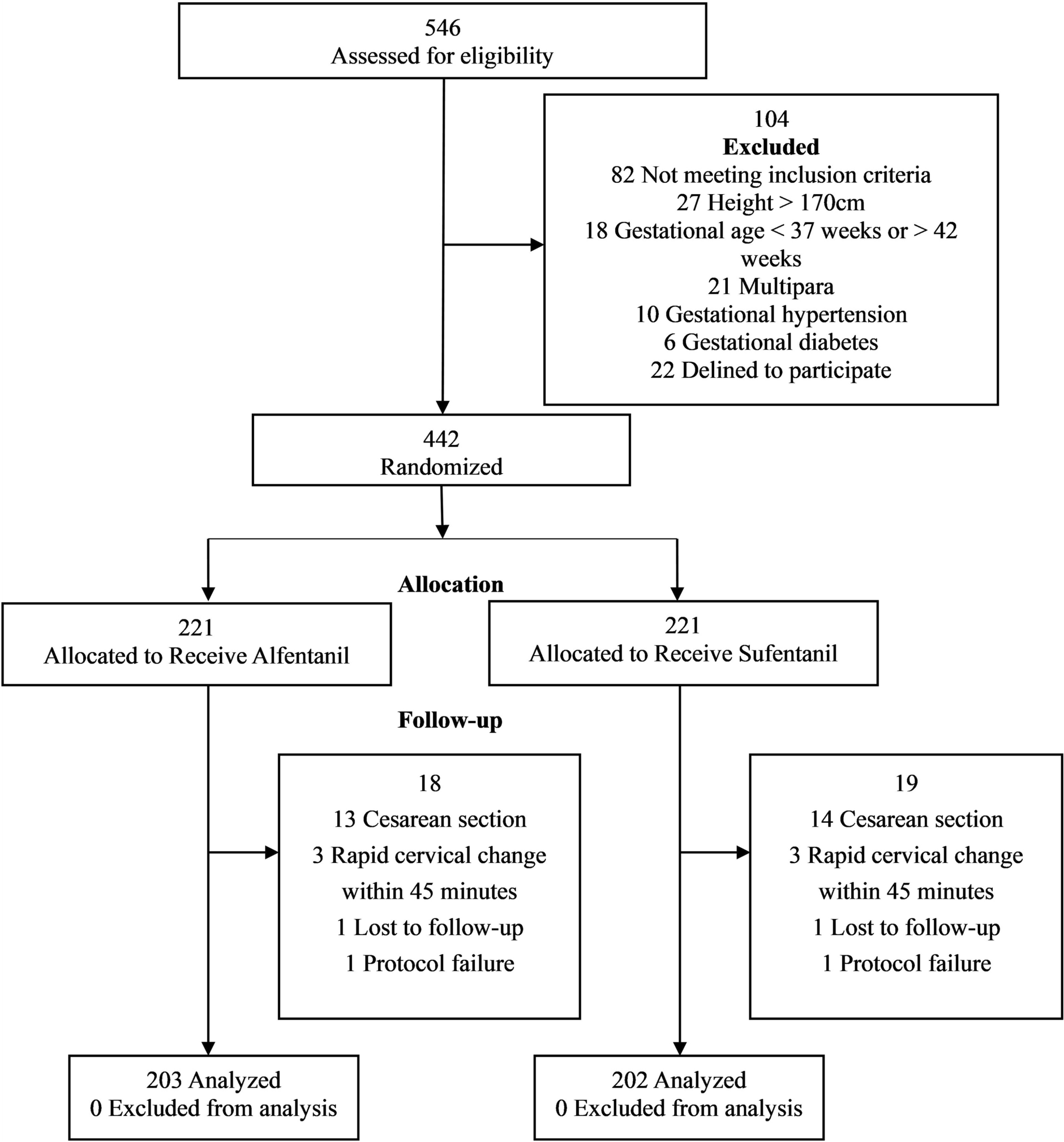
Other cells involved alongside T lymphocytes are B cells, which act as perpetuators of pain [54]. Their elimination using anti-CD20 therapy did not eliminate pain but merely delayed it, and at the end of the inflammatory cycle, B cells perpetuated it [55, 56]. Even with a low number of CD19+ B cells, pain was re-induced. In mouse models of chronic pain, B cells are activated and produce IgG; however, no specific autoantibody, like those found in encephalitis, has been identified. This seems to be a case of polyclonal activation triggered by uncontrolled innate immune responses at a specific site, leading to local antibody production and high expression of immunoglobulin receptors (FcγRs), and thus the perpetuation of inflammation and chronic pain [56]. The differentiation of naïve CD4+ T cells into distinct helper T cell subsets and their interactions with macrophages and B cells in modulating inflammation and pain are summarized in Fig. 2.
Fig. 2 The alt text for this image may have been generated using AI.
The alt text for this image may have been generated using AI.Key cellular mediators of chronic pain. Peripheral immune cells (macrophages, mast cells, T and B lymphocytes) and central glial cells (microglia, astrocytes) interact through cytokines, neurotrophic factors, and chemokines to drive peripheral and central sensitization. The schematic illustrates the differentiation of naïve CD4+ T cells into T helper (Th) 1, Th2, and Th17 subsets under specific polarizing cytokines, each producing distinct mediators that modulate immune responses. Pro-inflammatory subsets (Th1 and Th17) promote cytokine release and pain amplification, whereas anti-inflammatory Th2 cells contribute to immune regulation and pain resolution
Emerging Role of B Cells and Autoantibodies in Pain ChronificationRecent evidence has substantially expanded our understanding of B cell-mediated mechanisms in chronic pain beyond the polyclonal activation described previously. Lacagnina et al. [56] demonstrated that B cell depletion with anti-CD20 monoclonal antibody at the time of nerve injury prevented allodynia development in mice. Nerve injury induced immunoglobulin G (IgG) accumulation in ipsilateral lumbar DRG and dorsal spinal cords, where IgG colocalized with sensory neurons, macrophages, and microglia. The pathogenic role of autoantibodies extends across diverse conditions. In fibromyalgia, anti-satellite glial cell IgG antibodies correlate with pain severity, and passive transfer of patient IgG to mice induces pain-like behavior [57]. Similar findings have been reported in complex regional pain syndrome (CRPS), chronic post-traumatic limb pain, and rheumatoid arthritis-associated pain [58].
Multiple specific autoantibody targets have been identified. Autoantibodies against contactin-associated protein-like 2 (CASPR2), particularly IgG4 subclass, cause neuropathic pain by disrupting voltage-gated potassium channel function and inducing DRG hyperexcitability [59]. Autoantibodies against fibroblast growth factor receptor 3 (FGFR3) activate p38 MAPK, ERK, and JNK signaling pathways in sensory neurons [60]. Additional targets include plexin D1, anti-CV2/CRMP5, and novel antigens such as MX1, DBNL, and KRT8 [61]. These autoantibodies drive pain through multiple mechanisms: (1) complement system activation promoting inflammation and tissue damage; (2) signaling through Fc gamma receptors (FcγRs) on sensory neurons, triggering calcium mobilization via Syk and TRPC3 channels; (3) direct disruption of ion channel function; and (4) modulation of satellite glia and macrophage activity in DRG [28]. These mechanisms contribute to both peripheral and central sensitization. However, the predominance of IgG4 autoantibodies poses challenges, as IVIg contains minimal IgG4 and lacks sufficient idiotypic antibodies for neutralization, potentially explaining variable treatment responses. The identification of pain-sensitizing autoantibodies represents a paradigm shift in understanding chronic pain etiology, particularly in “unexplained” chronic primary pain conditions. These findings suggest that a subset of chronic pain patients may have an autoimmune basis, opening avenues for diagnostic biomarker development and targeted immunotherapies.
Pro-inflammatory versus Anti-inflammatory ResponsesCD8+ T lymphocytes also have dual roles: They can promote neuropathic pain by releasing pro-inflammatory cytokines, with cytotoxic CD8+ T cells primarily found in neuropathic pain associated with diabetes or HIV [62, 63]. However, they can also prevent the progression of neuropathic pain, as seen in a study of chemotherapy-induced neuropathic pain, where CD8+ T cells were observed to release IL-13, which in turn activates macrophages to secrete IL-10. This IL-10–rich microenvironment led to the resolution of mechanical allodynia [17].
During a certain phase of inflammation, everything is activation and pro-inflammatory signaling, with the release of cytokines. However, whether inflammation persists or resolves depends on the activation signals and the perpetuation of these signals in the dorsal horn of the spinal cord. For example, if the environment is rich in IL-12 and interferon-gamma (IFN-γ), a naïve T cell will differentiate into a Th1 cell; if the environment is rich in IL-6, IL-21, and transforming growth factor beta (TGF-β), it will become a Th17 cell; if the environment is rich in IL-4 and IL-2, it will become a Th2 cell; and if only TGF-β is present, it will differentiate into a regulatory T cell [48]. Each of these subtypes has different functions; the first two are pro-inflammatory, while the latter two are regulatory. Translated into pain mechanisms, if the DRG is infiltrated by Th1 and Th17 lymphocytes, macrophages, and neutrophils, this will lead to pain amplification and the onset of chronic pain [48]. Conversely, if the infiltrate consists of Th2 or regulatory T cells, the result will be the opposite: type 2 macrophages, pain resolution, and neural regeneration [48]. These represent two possible scenarios in the context of pain.
One of the main regulators of pain are regulatory T cells (CD4+CD25+), which are capable of secreting large amounts of IL-10, thereby suppressing the function of Th1 T lymphocytes. In this way, they help control inflammation and promote an anti-inflammatory state. This is one of the reasons that not all individuals develop chronic pain despite experiencing the same stimulus. The absence or failure to induce these regulatory cells can sometimes explain the persistence of chronic pain [26].
Influence of Local versus Systemic InflammationIL-10 has been demonstrated to exert antinociceptive effects when stimulated by other interleukins. For instance, intrathecal administration of IL-27 elicited IL-10-dependent antinociception in a peripheral neuropathic pain model. Likewise, intrathecal delivery of IL-35 reduced pain-related behaviors in experimental autoimmune encephalomyelitis and ameliorated diabetic neuropathic pain. Both interleukins enhance IL-10 production within inflamed environments [64, 65].
IL-10 expression occurs in monocytes, macrophages, dendritic cells, T cells, B cells, astrocytes, microglia, and oligodendrocytes, the very cells driving inflammation [66]. Insufficient IL-10 production has been linked to chronic pain, as IL-10 suppresses second messenger pathways that promote pro-inflammatory cytokine release [67]. Although its therapeutic application remains under debate, particularly concerning the optimal route of administration (local versus systemic) and dosage (low versus high), there is consensus that IL-10 is pivotal in regulating inflammation [68].
Inflammation is orchestrated by both adaptive and innate immunity. While T lymphocytes (T cells) are central to adaptive responses, innate cells, such as neutrophils and macrophages, are equally critical. These cells initiate and sustain inflammation but also contribute to its resolution and tissue repair, functions now recognized as integral to the inflammatory cascade. Once trauma subsides, mechanisms must actively suppress inflammation [69].
Peripheral monocytes infiltrate the DRG and differentiate into macrophages. Their function depends on the microenvironment, contributing initially to inflammation but later supporting resolution—an area still underexplored. Type 2 macrophages, induced by cytokines such as IL-4 and TGF-β, are central to neural repair and anti-inflammatory activity. They release endogenous opioids at injury sites, modulating local opioid signaling and pain perception. The absence of these cells may predispose individuals to chronic pain development [50].
Therapeutic Implications and InterventionsPolarization of macrophages toward the M2 phenotype and the consequent release of IL-10 have been shown to reduce inflammation and alleviate pain in murine models of osteoarthritis induced by platelet-rich plasma infiltration. In this context, reductions in IL-1 and IL-6 levels were accompanied by decreased expression of NGF, highlighting the close interplay between immune modulation and nociceptive signaling [70].
The immune system interacts extensively with both the central and peripheral nervous systems, primarily through microglia. These resident immune cells of the central nervous system share numerous surface markers with macrophages and monocytes. Neuropathic pain is accompanied by increased numbers and heightened activity of microglia; however, their inactivation after pain onset does not necessarily resolve symptoms [71]. Microglia secrete inflammatory mediators including colony-stimulating factor (CSF-1), neuregulin-1 (NRG-1), metalloproteases, and caspases, which activate Toll-like receptors (TLR2 and TLR4) on neutrophils. These neutrophils, in turn, amplify and perpetuate inflammation, releasing mediators that re-stimulate microglia and sustain a pathological feed-forward loop [72]. Notably, this process is sex-dependent: male mice recover more rapidly from neuropathic pain despite early surges of pro-inflammatory cytokines, likely reflecting more efficient nerve regeneration [73].
Although microglia were initially thought to induce chronic pain in concert with astrocytes [72, 74], subsequent studies demonstrated that their early inactivation or depletion, followed by repopulation, can paradoxically trigger the re-emergence of pain [9,
Comments (0)